Descifrarea genomului uman. Genetica medicală La descifrarea genomului Drosophila s-a constatat că
La 50 de ani de la descoperirea structurii ADN-ului
A.V. Zelenin
GENOMUL PLANTELOR
A. V. Zelenin
Zelenin Alexandru Vladimirovici- d.b.n.,
şef al laboratorului Institutului de Biologie Moleculară. V.A. Engelhardt RAS.
Realizările impresionante ale programului „Genom uman”, precum și succesul lucrărilor de descifrare a așa-numitelor genomuri extra-mici (virusuri), mici (bacterii, drojdie) și medii (viermi rotunzi, Drosophila), au făcut posibilă trecerea la un studiu pe scară largă a genomului plantelor mari și foarte mari. Necesitatea urgentă a unui studiu detaliat al genomilor celor mai importante plante din punct de vedere economic a fost subliniată la o întâlnire despre genomica plantelor, organizată în 1997 în Statele Unite [ , ]. De-a lungul anilor care au trecut de atunci, s-au obținut succese neîndoielnice în acest domeniu. În 2000, a apărut o publicație privind secvențierea completă (determinarea secvenței nucleotidice liniare a întregului ADN nuclear) a genomului muștarului mic - Arabidopsis, în 2001 - privind secvențierea preliminară (proiect) a genomului orezului. Au fost raportate în mod repetat lucrări de secvențiere a genomului plantelor mari și super-mari (porumb, secară, grâu), cu toate acestea, aceste rapoarte nu conțineau informații specifice și erau mai degrabă de natura declarațiilor de intenție.
Se presupune că decodificarea genomului plantelor va deschide perspective largi pentru știință și practică. În primul rând, identificarea de noi gene și lanțul de reglare genetică a acestora va crește semnificativ productivitatea plantelor prin utilizarea abordărilor biotehnologice. Odată cu descoperirea, izolarea, reproducerea (clonarea) și secvențierea genelor responsabile pentru funcții atât de importante ale organismului vegetal precum reproducerea și productivitatea, procesele de variabilitate, rezistența la factorii de mediu adversi, precum și împerecherea omoloagă a cromozomilor, apariția se asociază noi oportunităţi de îmbunătăţire a procesului de ameliorare . În cele din urmă, genele izolate și donate pot fi folosite pentru a obține plante transgenice cu proprietăți fundamental noi și pentru a analiza mecanismele de reglare a activității genelor.
Importanța studierii genomului plantelor este subliniată și de faptul că până în prezent numărul de gene de plante localizate, clonate și secvențiate este mic și variază, după diverse estimări, între 800 și 1200. Acesta este de 10-15 ori mai mic decât, pt. exemplu, la oameni.
Statele Unite rămân liderul incontestabil în studiul pe scară largă a genomului plantelor, deși studii intensive ale genomului orezului sunt efectuate în Japonia și în anul trecut si in China. La descifrarea genomului Arabidopsis, pe lângă laboratoarele din SUA, au participat activ grupurile de cercetare europene. Conducerea aparentă a Statelor Unite provoacă îngrijorare serioasă a oamenilor de știință europeni, pe care aceștia au exprimat-o în mod clar la o întâlnire sub titlul semnificativ „Perspective pentru genomica în era post-genomică”, desfășurată la sfârșitul anului 2000 în Franța. Progresul științei americane în studierea genomului plantelor agricole și crearea formelor de plante transgenice, potrivit oamenilor de știință europeni, amenință că într-un viitor nu prea îndepărtat (două-cinci decenii), când creșterea populației va pune omenirea în fața unui criza alimentară, economia și știința europeană vor deveni dependente de tehnologia americană. În acest sens, a fost anunțată crearea unui program științific franco-german pentru studiul genomului plantelor („Plantgene”) și s-au făcut investiții semnificative în acesta.
Evident, problemele genomicei plantelor ar trebui să atragă atenția oamenilor de știință și organizatorilor ruși ai științei, precum și a organelor de conducere, deoarece nu este vorba doar de prestigiul științific, ci și de securitatea națională a țării. Într-un deceniu sau două, alimentele vor deveni cea mai importantă resursă strategică.
DIFICULTĂȚI ÎN STUDIAREA GENOMULUI PLANTELOR
Studiul genomului plantelor este o sarcină mult mai dificilă decât studiul genomului oamenilor și al altor animale. Acest lucru se datorează următoarelor circumstanțe:
dimensiuni uriașe ale genomului, ajungând la anumite tipuri plante de zeci și chiar sute de miliarde de perechi de baze (pb): genoamele principalelor plante importante din punct de vedere economic (cu excepția orezului, inului și bumbacului) sunt fie apropiate ca mărime de genomul uman, fie de multe ori mai mari (tabel);STUDII CROMOZOMIALE ALE GENOMURILORFluctuații puternice ale numărului de cromozomi la diferite plante - de la două la unele specii la câteva sute la altele și nu este posibil să se identifice o corelație strictă între dimensiunea genomului și numărul de cromozomi;
Se formează o abundență de poliploide (conținând mai mult de doi genomi per celulă) cu genomi similari, dar nu identici (alpoliploidie);
Îmbogățirea extremă a genomului plantelor (până la 99%) de ADN „nesemnificativ” (necodificare, adică neconținând gene), ceea ce face foarte dificilă unirea (aranjarea în ordinea corectă) a fragmentelor secvențiate într-un mod comun. regiune ADN de dimensiuni mari (contig);
Cartografierea morfologică, genetică și fizică incompletă a cromozomilor (comparativ cu genomul Drosophila, uman și șoarece);
Imposibilitatea practică a izolării cromozomilor individuali în formă pură folosind metode utilizate de obicei în acest scop pentru cromozomi umani și animale (sortarea în flux și utilizarea hibrizilor celulari);
Dificultatea cartografierii cromozomiale (determinarea locației pe cromozom) a genelor individuale folosind hibridizare in situ, datorită atât conținutului ridicat de ADN „nesemnificativ” din genomul plantelor, cât și particularităților organizării structurale a cromozomilor plantelor;
Depărtarea evolutivă a plantelor față de animale, ceea ce complică serios utilizarea informațiilor obținute prin secvențierea genomului oamenilor și a altor animale pentru studiul genomului plantelor;
Procesul lung de reproducere al majorității plantelor, care încetinește semnificativ analiza genetică a acestora.
Studiile cromozomiale (citogenetice) ale genomului în general și ale plantelor în special au poveste lungă. Termenul „genom” a fost propus pentru a desemna un set haploid (unic) de cromozomi cu genele conținute în ei în primul sfert al secolului al XX-lea, adică cu mult înainte de stabilirea rolului ADN-ului ca purtător de informații genetice. .
Descrierea genomului unui organism multicelular nou, nestudiat genetic anterior, începe de obicei cu studiul și descrierea setului complet al cromozomilor săi (cariotip). Acest lucru, desigur, se aplică și plantelor, dintre care un număr mare nici măcar nu au început să fie studiate.
Deja în zorii studiilor cromozomiale, genomurile speciilor de plante înrudite au fost comparate pe baza analizei conjugării meiotice (combinație de cromozomi omologi) în hibrizi interspecifici. În ultimii 100 de ani, posibilitățile de analiză a cromozomilor s-au extins dramatic. Acum, pentru caracterizarea genomului plantelor sunt folosite tehnologii mai avansate: diverse variante ale așa-numitei colorări diferențiale, care face posibilă identificarea cromozomilor individuali după caracteristicile morfologice; hibridizare in situ făcând posibilă localizarea unor gene specifice pe cromozomi; cercetare biochimică proteine celulare (electroforeza si imunochimie) si, in final, un set de metode bazate pe analiza ADN-ului cromozomial pana la secventierea acestuia.

Orez. unu. Cariotipurile cerealelor a - secară (14 cromozomi), b - grâu dur (28 cromozomi), c - grâu moale (42 cromozomi), d - orz (14 cromozomi)De mulți ani, au fost studiate cariotipurile cerealelor, în primul rând grâul și secară. Este interesant că tipuri diferite aceste plante, numărul de cromozomi este diferit, dar întotdeauna un multiplu de șapte. Tipurile individuale de cereale pot fi recunoscute în mod fiabil după cariotipul lor. De exemplu, genomul de secară este format din șapte perechi de cromozomi mari cu blocuri heterocromatice intens colorate la capete, adesea numite segmente sau benzi (Fig. 1a). Genomul grâului are deja 14 și 21 de perechi de cromozomi (Fig. 1, b, c), iar distribuția blocurilor heterocromatice în ele nu este aceeași ca și în cromozomii de secară. Genomii individuali de grâu, desemnați A, B și D, diferă, de asemenea, unul de celălalt. O creștere a numărului de cromozomi de la 14 la 21 duce la o schimbare bruscă a proprietăților grâului, care se reflectă în numele lor: durum, sau paste, grâu și moale, sau pâine, grâu . Gena D, care conține gene pentru proteinele glutenului, care conferă aluatului așa-numita germinație, este responsabilă pentru dobândirea de înalte proprietăți de coacere de către grâul moale. Acest genom este căruia i se acordă o atenție deosebită în îmbunătățirea selecției grâului de pâine. O altă cereală cu 14 cromozomi, orzul (Fig. 1, d), nu este de obicei folosită pentru a face pâine, dar este principala materie primă pentru fabricarea unor produse atât de comune precum berea și whisky-ul.
Cromozomii unor plante sălbatice utilizate pentru îmbunătățirea calității celor mai importante specii agricole, precum rudele sălbatice ale grâului - Aegilops, sunt în curs de studiu intens. Noi forme de plante sunt create prin încrucișare (Fig. 2) și selecție. În ultimii ani, o îmbunătățire semnificativă a metodelor de cercetare a făcut posibilă începerea studiului genomului plantelor, ale căror cariotipuri (în principal dimensiunea mică a cromozomilor) le făceau anterior inaccesibile pentru analiza cromozomilor. Așadar, abia recent au fost identificați pentru prima dată toți cromozomii de bumbac, mușețel și in.

Orez. 2. Cariotipuri de grâu și un hibrid de grâu cu Aegilops
a - grâu moale hexaploid ( Triticum astivum), constând din genomi A, B și O; b - grâu tetraploid ( Triticum timopheevi), constând din genomi A și G. conține gene de rezistență la majoritatea bolilor grâului; c - hibrizi Triticum astivum X Triticum timopheevi rezistent la mucegai și rugină, înlocuirea unei părți a cromozomilor este clar vizibilăSTRUCTURA PRIMARĂ A ADN-ului
Odată cu dezvoltarea geneticii moleculare, însuși conceptul de genom s-a extins. Acum acest termen este interpretat atât în sensul cromozomial clasic, cât și în sensul molecular modern: întregul material genetic al unui virus, al unei celule și al unui organism individual. Desigur, în urma studiului structurii primare complete a genomului (cum este adesea numită secvența liniară completă a bazelor de acid nucleic) a unui număr de microorganisme și oameni, a apărut problema secvențierii genomului plantelor.
Dintre numeroasele organisme vegetale, două au fost selectate pentru studiu - Arabidopsis, reprezentând clasa dicotiledoneelor (dimensiunea genomului 125 milioane bp) și orez din clasa monocotiledonelor (420-470 milioane bp). Acești genomi sunt mici în comparație cu alți genomi de plante și conțin relativ puține segmente ADN repetitive. Astfel de caracteristici au dat speranța că genomii selectați vor fi disponibili pentru determinarea relativ rapidă a structurii lor primare.

Orez. 3. Arabidopsis - muștar mic - o plantă mică din familia cruciferelor ( Brassicaceae). Pe un spațiu egal ca suprafață cu o pagină a revistei noastre, puteți crește până la o mie de organisme Arabidopsis individuale.Motivul alegerii Arabidopsis nu a fost doar dimensiunea redusă a genomului său, ci și dimensiunea redusă a organismului, ceea ce face ușoară creșterea lui în laborator (Fig. 3). Am luat în considerare ciclul său de reproducere scurt, datorită căruia este posibil să se efectueze rapid experimente privind încrucișarea și selecția, genetica studiată în detaliu, ușurința de manipulare în condițiile de creștere în schimbare (schimbarea compoziției de sare a solului, adăugarea diverșilor nutrienți etc. .) și testarea efectului asupra plantelor a diverșilor factori mutageni și agenți patogeni (viruși, bacterii, ciuperci). Arabidopsis nu are valoare economică, prin urmare, genomul său, împreună cu genomul șoarecelui, a fost numit referință sau, mai puțin precis, model.*
* Apariția termenului „genom model” în literatura rusă este rezultatul unei traduceri inexacte a expresiei engleze model genom. Cuvântul „model” înseamnă nu numai adjectivul „model”, ci și substantivul „probă”, „standard”, „model”. Ar fi mai corect să vorbim despre un genom eșantion sau despre un genom de referință.Lucrările intensive privind secvențierea genomului Arabidopsis au fost începute în 1996 de un consorțiu internațional care includea instituții științifice și grupuri de cercetare din SUA, Japonia, Belgia, Italia, Marea Britanie și Germania. În decembrie 2000, au devenit disponibile informații extinse care rezumă determinarea structurii primare a genomului Arabidopsis. Pentru secvențiere a fost folosită tehnologia clasică sau ierarhică: mai întâi, au fost studiate secțiuni individuale mici ale genomului, din care au fost compuse secțiuni mai mari (contigs) și, în stadiul final, structura cromozomilor individuali. ADN-ul nuclear al genomului Arabidopsis este distribuit pe cinci cromozomi. În 1999, au fost publicate rezultatele secvențierii a doi cromozomi, iar apariția în presă a informațiilor despre structura primară a celor trei rămași a completat secvențierea întregului genom.
Din 125 de milioane de perechi de baze, a fost determinată structura primară de 119 milioane, ceea ce reprezintă 92% din întregul genom. Doar 8% din genomul Arabidopsis care conține blocuri mari de segmente repetitive de ADN s-a dovedit a fi inaccesibil pentru studiu. În ceea ce privește completitatea și minuțiozitatea secvențierii genomului eucariotic, Arabidopsis rămâne în primii trei campioni împreună cu un organism de drojdie unicelulară. Saccharomyces cerevisiaeși organism multicelular Caenorhabditis eleganta(Vezi tabelul).
Aproximativ 15.000 de gene individuale care codifică proteine au fost găsite în genomul Arabidopsis. Aproximativ 12.000 dintre acestea sunt conținute sub formă de două copii per genom haploid (unic), astfel încât numărul total genele este de 27 mii. Numărul de gene din Arabidopsis nu diferă mult de numărul de gene din organisme precum oamenii și șoarecii, dar dimensiunea genomului său este de 25-30 de ori mai mică. Această circumstanță este asociată cu caracteristici importante în structura genelor individuale Arabidopsis și structura generală a genomului său.
Genele Arabidopsis sunt compacte, conținând doar câțiva exoni (regiuni care codifică proteine) separați de segmente scurte (aproximativ 250 bp) de ADN necodificatori (introni). Intervalele dintre genele individuale sunt în medie de 4600 de perechi de baze. Pentru comparație, subliniem că genele umane conțin multe zeci și chiar sute de exoni și introni, iar regiunile intergenice au dimensiuni de 10 mii de perechi de baze sau mai mult. Se presupune că prezența unui genom compact mic a contribuit la stabilitatea evolutivă a Arabidopsis, deoarece ADN-ul său a devenit o țintă pentru diferiți agenți dăunători într-o măsură mai mică, în special pentru introducerea de fragmente de ADN repetitive asemănătoare virusului (transpozoni). în genom.
Printre alte caracteristici moleculare ale genomului Arabidopsis, trebuie remarcat faptul că exonii sunt îmbogățiți în guanină și citozină (44% în exoni și 32% în introni) în comparație cu genele animale, precum și prezența genelor dublu repetate (duplicate). Se presupune că o astfel de dublare a avut loc ca urmare a a patru evenimente simultane, constând în dublarea (repetarea) unei părți a genelor Arabidopsis sau fuziunea genomilor înrudite. Aceste evenimente, care au avut loc în urmă cu 100-200 de milioane de ani, sunt o manifestare a tendinței generale de poliploidizare (o creștere multiplă a numărului de genomi dintr-un organism), care este caracteristică genomului plantelor. Cu toate acestea, unele fapte arată că genele duplicate în Arabidopsis nu sunt identice și funcționează diferit, ceea ce poate fi asociat cu mutații în regiunile lor de reglementare.
Orezul a devenit un alt obiect de secvențiere completă a ADN-ului. Genomul acestei plante este, de asemenea, mic (12 cromozomi, dând un total de 420-470 milioane bp), doar de 3,5 ori mai mare decât cel al Arabidopsis. Cu toate acestea, spre deosebire de Arabidopsis, orezul are o mare importanță economică, fiind baza de nutriție pentru mai mult de jumătate din umanitate, prin urmare, nu numai miliarde de consumatori, ci și o armată de oameni multimilionare implicați activ în procesul foarte laborios al acestuia. cultivarea sunt extrem de interesați să-și îmbunătățească proprietățile.
Unii cercetători au început să studieze genomul orezului încă din anii 1980, dar aceste studii au atins o amploare serioasă abia în anii 1990. În 1991, în Japonia a fost creat un program de descifrare a structurii genomului orezului, reunind eforturile multor grupuri de cercetare. În 1997, pe baza acestui program a fost organizat Proiectul Internațional al Genomului Orezului. Participanții săi au decis să-și concentreze eforturile pe secvențierea uneia dintre subspeciile de orez ( Oriza sativajaponica), în studiul căruia s-au înregistrat deja progrese semnificative până la acel moment. Un stimulent serios și, la figurat vorbind, o vedetă călăuzitoare pentru o astfel de muncă a fost programul „Genom uman”.
În cadrul acestui program a fost testată strategia de diviziune ierarhică „cromozomială” a genomului, pe care participanții consorțiului internațional au folosit-o pentru a descifra genomul orezului. Cu toate acestea, dacă în studiul genomului uman, fracțiile de cromozomi individuali au fost izolate folosind diverse metode, atunci materialul specific pentru cromozomii individuali de orez și regiunile lor individuale a fost obținut prin microdisecție cu laser (decuparea obiectelor microscopice). Pe o lamă de microscop, unde se află cromozomii de orez, sub influența unui fascicul laser, totul este ars, cu excepția cromozomului sau a secțiunilor acestuia programate pentru analiză. Materialul rămas este folosit pentru clonare și secvențiere.
Au fost publicate numeroase rapoarte cu privire la rezultatele secvențierii fragmentelor individuale ale genomului orezului, efectuate cu mare acuratețe și detaliu, caracteristice tehnologiei ierarhice. Se credea că determinarea structurii primare complete a genomului orezului va fi finalizată până la sfârșitul anului 2003 – mijlocul anului 2004, iar rezultatele, împreună cu datele privind structura primară a genomului Arabidopsis, vor fi utilizate pe scară largă în comparație. genomica altor plante.
Cu toate acestea, la începutul anului 2002, două grupuri de cercetare - unul din China, celălalt din Elveția și Statele Unite - au publicat rezultatele unui proiect complet de secvențiere (aproximativă) a genomului orezului, realizat folosind tehnologia de clonare totală. Spre deosebire de studiul în etape (ierarhice), abordarea totală se bazează pe clonarea simultană a întregului ADN genomic într-unul dintre vectorii virali sau bacterieni și obținerea unui număr semnificativ (uriaș pentru genomi medii și mari) de clone individuale care conțin diverse Segmente de ADN. Pe baza analizei acestor secțiuni secvențiate și a suprapunerii secțiunilor terminale identice ale ADN-ului, se formează un contig - un lanț de secvențe ADN unite între ele. Contigul general (total) este structura primară a întregului genom sau cel puțin a unui cromozom individual.
Într-o astfel de prezentare schematică, strategia clonării totale pare simplă. De fapt, întâmpină dificultăți serioase asociate cu necesitatea obținerii unui număr imens de clone (se acceptă în general că genomul sau regiunea sa studiată trebuie să fie suprapuse de clone de cel puțin 10 ori), o cantitate imensă de secvențiere și extrem de complexă. munca la clone de andocare care necesită participarea specialiștilor în bioinformatică. Un obstacol serios în calea clonării totale este o varietate de segmente ADN repetitive, al căror număr, după cum sa menționat deja, crește brusc pe măsură ce dimensiunea genomului crește. Prin urmare, strategia de secvențiere totală este utilizată în principal în studiul genomului virusurilor și microorganismelor, deși a fost folosită cu succes pentru a studia genomul unui organism multicelular, Drosophila.
Rezultatele secvențierii totale a acestui genom au fost „suprapuse” unei game uriașe de informații despre structura sa cromozomială, genică și moleculară, obținute pe o perioadă de aproape 100 de ani de studiu a Drosophila. Și totuși, în ceea ce privește gradul de secvențiere, genomul Drosophila (66% din dimensiunea totală a genomului) este semnificativ inferior genomul Arabidopsis (92%), în ciuda dimensiunilor lor destul de apropiate - 180 de milioane și, respectiv, 125 de milioane de perechi de baze. . Prin urmare, recent s-a propus denumirea tehnologiei mixte, care a fost folosită pentru secvențierea genomului Drosophila.
Pentru a secvenționa genomul orezului, grupurile de cercetare menționate mai sus au luat două dintre subspeciile sale, cele mai cultivate în țările asiatice, - Oriza saliva L. ssp indicajși Oriza saliva L. sspjaponica. Rezultatele studiilor lor coincid în multe privințe, dar diferă în multe privințe. Astfel, reprezentanții ambelor grupuri au declarat că au ajuns la aproximativ 92-93% din suprapunerea genomului cu contigs. S-a demonstrat că aproximativ 42% din genomul orezului este reprezentat de repetări scurte de ADN constând din 20 de perechi de baze, iar majoritatea elementelor mobile de ADN (transpozoni) sunt localizate în regiuni intergenice. Cu toate acestea, datele privind dimensiunea genomului orezului diferă semnificativ.
Pentru subspecia japoneză, dimensiunea genomului este determinată a fi de 466 milioane de perechi de baze, iar pentru subspecia indiană, 420 milioane. Motivul acestei discrepanțe nu este clar. Poate fi o consecință a diferitelor abordări metodologice în determinarea dimensiunii părții necodificatoare a genomului, adică nu reflectă adevărata stare a lucrurilor. Dar este posibil să existe o diferență de 15% în dimensiunea genomilor studiati.
A doua discrepanță majoră a fost dezvăluită în numărul de gene găsite: pentru subspecia japoneză, de la 46 022 la 55 615 gene per genom, iar pentru subspecia indiană, de la 32 000 la 50 000. Motivul acestei discrepanțe nu este clar.
Incompletitudinea și inconsecvența informațiilor primite se remarcă în comentariile la articolele publicate. De asemenea, aici se exprimă speranța că lacunele în cunoașterea genomului orezului vor fi eliminate prin compararea datelor de „secvențiere brută” cu rezultatele secvențierii detaliate, ierarhice, efectuate de participanții la Proiectul Internațional de Genom al Orezului.
GENOMIA PLANTELOR COMPARATIVA SI FUNCTIONALA
Datele extinse obținute, dintre care jumătate (rezultatele grupului chinez) sunt disponibile publicului, deschid fără îndoială perspective largi atât pentru studiul genomului orezului, cât și pentru genomica plantelor în general. O comparație a proprietăților genomului Arabidopsis și al orezului a arătat că majoritatea genelor (până la 80%) identificate în genomul Arabidopsis se găsesc și în genomul orezului, totuși, pentru aproximativ jumătate din genele găsite în orez, analogi (ortologi) ) nu au fost încă găsite în genomul Arabidopsis. În același timp, 98% dintre genele a căror structură primară a fost stabilită pentru alte cereale au fost găsite în genomul orezului.
Discrepanța semnificativă (aproape dublă) dintre numărul de gene din orez și Arabidopsis este uluitoare. În același timp, datele proiectului de decodificare a genomului orezului, obținute prin secvențierea totală, practic nu sunt comparate cu rezultatele ample ale studiului genomului orezului prin metoda clonării și secvențierii ierarhice, adică ceea ce are nu a fost efectuată cu privire la genomul Drosophila. Prin urmare, rămâne neclar dacă diferența în numărul de gene în Arabidopsis și orez reflectă adevărata stare de lucruri sau dacă este explicată prin diferența de abordări metodologice.
Spre deosebire de genomul Arabidopsis, nu sunt date date despre genele gemene din genomul orezului. Este posibil ca cantitatea lor relativă să fie mai mare în orez decât în Arabidopsis. Această posibilitate este susținută indirect de date privind prezența formelor poliploide de orez. Se poate aștepta mai multă claritate cu privire la această problemă după finalizarea Proiectului Internațional de Genom al Orezului și se obține o imagine detaliată a structurii primare a ADN-ului acestui genom. Motive serioase pentru o astfel de speranță sunt oferite de faptul că, după publicarea lucrărilor privind secvențierea brută a genomului de orez, numărul de publicații privind structura acestui genom a crescut brusc, în special, au apărut informații despre secvențierea detaliată. dintre cromozomii 1 și 4 ai săi.
Cunoașterea, cel puțin aproximativ, a numărului de gene din plante este de o importanță fundamentală pentru genomica comparativă a plantelor. Inițial, s-a crezut că, deoarece toate plantele cu flori sunt foarte apropiate una de cealaltă în ceea ce privește caracteristicile lor fenotipice, genomul lor ar trebui să fie, de asemenea, similar. Și dacă studiem genomul Arabidopsis, vom obține informații despre majoritatea genomilor altor plante. O confirmare indirectă a acestei presupuneri sunt rezultatele secvențierii genomului șoarecelui, care este surprinzător de aproape de genomul uman (aproximativ 30 de mii de gene, dintre care doar 1 mie s-au dovedit a fi diferite).
Se poate presupune că motivul diferențelor dintre genomul Arabidopsis și orezul constă în apartenența lor la diferite clase de plante - dicotiledone și monocotiledone. Pentru a clarifica această problemă, este foarte de dorit să se cunoască cel puțin o structură primară brută a unei alte plante monocotiledonate. Cel mai realist candidat ar putea fi porumbul, al cărui genom este aproximativ egal cu genomul uman, dar totuși mult mai mic decât genomul altor cereale. Valoarea nutritivă a porumbului este binecunoscută.
Vastul material obținut ca urmare a secvențialării genomului Arabidopsis și orezului devine treptat baza unui studiu la scară largă al genomului plantelor folosind genomica comparativă. Astfel de studii au o semnificație biologică generală, deoarece ne permit să stabilim principiile principale de organizare a genomului plantei în ansamblu și cromozomii lor individuali, pentru a identifica aspecte comune structura genelor și regiunile lor de reglare, pentru a lua în considerare raportul dintre partea activă funcțional (genă) a cromozomului și diferitele regiuni ADN intergenice care nu codifică proteine. Genetica comparativă devine, de asemenea, din ce în ce mai importantă pentru dezvoltarea genomicii funcționale umane. Pentru studii comparative s-a efectuat secvențierea genomilor de pește-papă și șoarece.
La fel de important este studiul genelor individuale responsabile de sinteza proteinelor individuale care determină funcțiile specifice ale corpului. Este în descoperirea, izolarea, secvențierea și determinarea funcției genelor individuale în care se află semnificația practică, în primul rând medicală, a programului Genomului uman. Această împrejurare a fost remarcată în urmă cu câțiva ani de către J. Watson, care a subliniat că programul „Genom uman” va fi finalizat numai atunci când funcțiile tuturor genelor umane vor fi determinate.

Orez. patru. Clasificare în funcție de funcția genelor Arabidopsis
1 - gene pentru creștere, diviziune și sinteza ADN; 2 - gene de sinteză a ARN (transcripție); 3 - gene pentru sinteza si modificarea proteinelor; 4 - gene pentru dezvoltare, îmbătrânire și moarte celulară; 5 - gene ale metabolismului celular și ale metabolismului energetic; 6 - gene de interacțiune intercelulară și transmitere a semnalului; 7 - gene pentru furnizarea altor procese celulare; 8 - gene cu funcție necunoscutăÎn ceea ce privește funcția genelor plantelor, știm mai puțin de o zecime din ceea ce știm despre genele umane. Chiar și în Arabidopsis, al cărui genom este mult mai studiat decât genomul uman, funcția a aproape jumătate din genele sale rămâne necunoscută (Fig. 4). Între timp, pe lângă genele comune animalelor, plantele au un număr semnificativ de gene care sunt specifice doar (sau cel puțin predominant) pentru ele. Este despre despre genele implicate în transportul apei și sinteza peretelui celular, care este absent la animale, despre genele care asigură formarea și funcționarea cloroplastelor, fotosinteza, fixarea azotului și sinteza a numeroase produse aromatice. Această listă poate fi continuată, dar este deja clar cu ce sarcină dificilă se confruntă genomica funcțională a plantelor.
Secvențierea întregului genom oferă informații aproape adevărate despre numărul total de gene dintr-un organism dat, face posibilă plasarea unor informații mai mult sau mai puțin detaliate și fiabile despre structura lor în băncile de date și facilitează munca de izolare și studiere a genelor individuale. Cu toate acestea, secvențierea genomului nu înseamnă în niciun caz să stabilească funcția tuturor genelor.
Una dintre cele mai promițătoare abordări ale genomicii funcționale se bazează pe identificarea genelor de lucru care sunt utilizate pentru transcrierea (citirea) ARNm. Această abordare, inclusiv tehnologie moderna microarrays, vă permite să identificați simultan până la zeci de mii de gene funcționale. Recent, folosind această abordare, a început studiul genomului plantelor. Pentru Arabidopsis, a fost posibil să se obțină aproximativ 26 de mii de transcrieri individuale, ceea ce facilitează foarte mult posibilitatea de a determina funcția aproape tuturor genelor sale. La cartofi, a fost posibil să se identifice aproximativ 20.000 de gene de lucru care sunt importante pentru înțelegerea atât a proceselor de creștere și formare a tuberculilor, cât și a proceselor de boală a cartofului. Se presupune că aceste cunoștințe vor crește rezistența unuia dintre cele mai importante produse alimentare la agenți patogeni.
Dezvoltarea logică a genomicii funcționale a fost proteomica. Acest nou domeniu de știință studiază proteomul, care este de obicei înțeles ca un set complet de proteine dintr-o celulă la un anumit moment. Un astfel de set de proteine, care reflectă starea funcțională a genomului, se schimbă tot timpul, în timp ce genomul rămâne neschimbat.
Studiul proteinelor a fost folosit de mult timp pentru a judeca activitatea genomului plantelor. După cum se știe, enzimele prezente în toate plantele diferă în fiecare specie și soiuri în secvența de aminoacizi. Astfel de enzime, cu aceeași funcție, dar cu o secvență diferită de aminoacizi individuali, se numesc izoenzime. Au proprietăți fizico-chimice și imunologice diferite (greutate moleculară, sarcină), care pot fi detectate prin cromatografie sau electroforeză. De mulți ani, aceste metode au fost folosite cu succes pentru a studia așa-numitul polimorfism genetic, adică diferențele dintre organisme, soiuri, populații, specii, în special grâul și formele înrudite de cereale. Recent, însă, datorită dezvoltării rapide a metodelor de analiză a ADN-ului, inclusiv secvențierea, studiul polimorfismului proteic a fost înlocuit cu studiul polimorfismului ADN. Cu toate acestea, studiul direct al spectrelor proteinelor de depozitare (prolamine, gliadine etc.), care determină principalele proprietăți nutriționale ale cerealelor, rămâne o metodă importantă și de încredere pentru analiza genetică, selecția și producția de semințe a plantelor agricole.
Cunoașterea genelor, a mecanismelor de exprimare și reglare a acestora este extrem de importantă pentru dezvoltarea biotehnologiei și producerea de plante transgenice. Se știe că succesele impresionante în acest domeniu provoacă o reacție ambiguă din partea comunității de mediu și medicale. Cu toate acestea, există o zonă a biotehnologiei vegetale în care aceste temeri, dacă nu complet nefondate, atunci, în orice caz, par a fi de puțină importanță. Vorbim despre crearea de plante industriale transgenice care nu sunt folosite ca produse alimentare. India a recoltat recent prima recoltă de bumbac transgenic care este rezistent la o serie de boli. Există informații despre introducerea unor gene speciale care codifică proteine pigmentare în genomul bumbacului și producerea de fibre de bumbac care nu necesită vopsire artificială. O altă cultură industrială care poate face obiectul unei inginerie genetică eficientă este inul. Utilizarea sa ca alternativă la bumbac pentru materii prime textile a fost discutată recent. Această problemă este extrem de importantă pentru țara noastră, care și-a pierdut propriile surse de bumbac brut.
PERSPECTIVE DE STUDIAREA GENOMULUI PLANTELOR
Evident, studiile structurale ale genomului plantelor se vor baza pe abordările și metodele genomicei comparative, folosind ca material principal rezultatele descifrării genomului Arabidopsis și orezului. Un rol important în dezvoltarea genomicii comparative a plantelor îl vor juca, fără îndoială, informațiile care mai devreme sau mai târziu vor fi furnizate de secvențierea totală (aproximată) a genomilor altor plante. În acest caz, genomica comparativă a plantelor se va baza pe stabilirea de relații genetice între loci individuali și cromozomi aparținând unor genomi diferiți. Ne vom concentra nu atât pe genomica generală a plantelor, cât pe genomica selectivă a locurilor cromozomiale individuale. De exemplu, s-a demonstrat recent că gena responsabilă pentru vernalizare este localizată la locusul VRn-AI al cromozomului hexaploid de grâu 5A și locusul Hd-6 al cromozomului 3 de orez.
Dezvoltarea acestor studii va fi un impuls puternic pentru identificarea, izolarea și secvențierea multor gene ale plantelor importante din punct de vedere funcțional, în special, genele responsabile de rezistența la boli, rezistența la secetă și adaptabilitatea la diferite condiții de creștere. Din ce în ce mai mult, se va folosi genomica funcțională, bazată pe detectarea în masă (screening) a genelor care funcționează în plante.
Putem prevedea îmbunătățiri suplimentare ale tehnologiilor cromozomiale, în primul rând metoda de microdisecție. Utilizarea sa extinde dramatic posibilitățile cercetării genomice fără a necesita costuri uriașe, cum ar fi, de exemplu, secvențierea totală a genomului. Metoda de localizare pe cromozomii plantelor a genelor individuale cu ajutorul hibridizării va fi răspândită în continuare. in situ.În prezent, utilizarea sa este limitată de numărul mare de secvențe repetitive din genomul plantei și, eventual, de particularitățile organizării structurale a cromozomilor plantelor.
Tehnologiile cromozomiale vor deveni de mare importanță pentru genomica evolutivă a plantelor în viitorul apropiat. Aceste tehnologii relativ ieftine fac posibilă evaluarea rapidă a variabilității intra și interspecifice, studiul genomilor alopoliploizi complexe de grâu tetraploid și hexaploid, triticale; analiza proceselor evolutive la nivel cromozomial; investighează formarea genomilor sintetici și introducerea (introgresiunea) materialului genetic străin; identificarea relațiilor genetice dintre cromozomii individuali diferite feluri.
Studiul cariotipului plantelor folosind metode citogenetice clasice, îmbogățit prin analiză biologică moleculară și tehnologie computerizată, va fi utilizat pentru caracterizarea genomului. Acest lucru este deosebit de important pentru studierea stabilității și variabilității cariotipului la nivelul nu numai al organismelor individuale, ci și al populațiilor, soiurilor și speciilor. În cele din urmă, este dificil de imaginat cum poate fi estimat numărul și spectrele de rearanjamente cromozomiale (aberații, punți) fără utilizarea metodelor de colorare diferențială. Astfel de studii sunt extrem de promițătoare pentru monitorizarea mediului în funcție de starea genomului plantei.
LA Rusia modernă secvențierea directă a genomului plantelor este puțin probabilă. O astfel de muncă, care necesită investiții mari, depășește puterea economiei noastre actuale. Între timp, datele privind structura genomului Arabidopsis și orezului, obținute de știința mondială și disponibile în băncile internaționale de date, sunt suficiente pentru dezvoltarea genomicii plantelor interne. Se poate prevedea extinderea studiilor asupra genomului plantelor bazate pe abordări genomice comparative pentru a rezolva probleme specifice de ameliorare și producție a culturilor, precum și pentru a studia originea diferitelor specii de plante de mare importanță economică.
Se poate presupune că abordările genomice precum tiparea genetică (analize RELF, RAPD, AFLP etc.), care sunt destul de accesibile pentru bugetul nostru, vor fi utilizate pe scară largă în practica internă de ameliorare și producția de culturi. În paralel cu metodele directe de determinare a polimorfismului ADN, abordări bazate pe studiul polimorfismului proteic, în primul rând proteinele de depozitare a cerealelor, vor fi utilizate pentru rezolvarea problemelor de genetică și ameliorare a plantelor. Tehnologiile cromozomiale vor fi utilizate pe scară largă. Sunt relativ ieftine, dezvoltarea lor necesită investiții destul de moderate. În domeniul studiilor cromozomiale, știința domestică nu este inferioară lumii.
Trebuie subliniat faptul că știința noastră a adus o contribuție semnificativă la formarea și dezvoltarea genomicii plantelor [ , ].
Rolul fundamental l-a jucat N.I. Vavilov (1887-1943).
În biologia moleculară și genomica plantelor, contribuția de pionierat a lui A.N. Belozerski (1905-1972).
În domeniul studiilor cromozomiale, este de remarcat munca remarcabilului genetician S.G. Navashin (1857-1930), care a descoperit primul cromozomi sateliti la plante și a demonstrat că este posibil să se facă distincția între cromozomi individuali în funcție de caracteristicile morfologiei lor.
Un alt clasic stiinta ruseasca G.A. Levitsky (1878-1942) a descris în detaliu cromozomii de secară, grâu, orz, mazăre și sfeclă de zahăr, a introdus termenul „cariotip” în știință și a dezvoltat doctrina acestuia.
Specialiștii moderni, bazându-se pe realizările științei mondiale, pot aduce o contribuție semnificativă la dezvoltarea ulterioară a geneticii și genomicii plantelor.
Autorul își exprimă sincere mulțumiri academicianului Yu.P. Altukhov pentru discuția critică a articolului și sfaturi valoroase.Activitatea echipei conduse de autorul articolului a fost susținută de Fundația Rusă pentru Cercetare de bază (granturi nr. 99-04-48832; 00-04-49036; 00-04-81086), Federația Rusă pentru sprijinirea școlilor științifice (granturi Nr. 00-115-97833 și NSh-1794.2003.4) și Programul Academia RusăȘtiințe „Markeri genetici moleculari și cromozomiali în dezvoltarea metodelor moderne de ameliorare și producție de semințe”.
LITERATURĂ
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Introducere în genomica plantelor // Biologie moleculară. 2001. V. 35. S. 339-348. 2. Pen E. Bonanza pentru genomica plantelor // Știință. 1998. V. 282. P. 652-654. 3. Genomica plantelor, Proc. Natl. Acad. sci. STATELE UNITE ALE AMERICII. 1998. V. 95. P. 1962-2032. 4. Cartel N.A. si etc. Genetica. Dicţionar enciclopedic. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferențierea genomului în Aegilops. 1. Distribuția secvențelor de ADN foarte repetitive pe cromozomii speciilor diploide, Genomul. 1996. V. 39. P. 293-306. Istoricul analizei cromozomilor // Biol. membranelor. 2001. T. 18. S. 164-172.
Pe 09.05.2011 la 09:36, Limarev a spus:
Limarev V.N.
Descifrarea genomului uman.
Fragment din cartea lui L.G. Puchko: „Cunoașterea radietică a omului”
Pentru a rezolva problemele de descifrare a genomului, proiectul internațional „Genom uman” a fost organizat cu un buget de miliarde de dolari.
Până în 2000, harta genomului uman era practic completă. Genele au fost numărate, identificate și înregistrate în baze de date. Acestea sunt cantități uriașe de informații.
Înregistrarea genomului uman în formă digitalizată necesită aproximativ 300 de terabytes de memorie de computer, ceea ce echivalează cu 3.000 de hard disk-uri cu o capacitate de 100 de gigabytes.
S-a dovedit. Că o persoană nu are sute de mii, așa cum se credea anterior, ci puțin mai mult de 30 de mii de gene. Musca are Drosophila, sunt doar jumătate din câte - aproximativ 13 mii, iar șoarecele are aproape același număr ca o persoană. Genele unice pentru oameni în genomul decodificat sunt doar aproximativ 1%. Cea mai mare parte a spiralei ADN, după cum s-a dovedit, este ocupată nu de gene, ci de așa-numitele „secțiuni goale”, în care genele pur și simplu nu sunt codificate, precum și fragmente duble care se repetă unul după altul, sensul și sensul. despre care este neclar.
Într-un cuvânt, genele s-au dovedit a fi nici măcar cărămizile vieții, ci doar elemente ale planului, conform cărora este construită clădirea organismului. Cărămizile, ca și în alte lucruri se credea înainte de perioada de glorie a geneticii, sunt proteine.
A devenit absolut evident că în 1% dintre genele unice pentru oameni, o cantitate atât de mare de informații care distinge oamenii de șoareci nu pot fi codificate. Unde sunt stocate toate informațiile? Pentru mulți oameni de știință, devine un fapt neîndoielnic că fără principiul divin este imposibil de explicat natura omului. O serie de oameni de știință sugerează că, în cadrul ideilor existente despre corpul uman, este practic imposibil să se descifreze genomul uman.
Lumea nu este cunoscută - este cunoscută (comentariile mele la articol).
1) Luați în considerare fragmentul: „Fără principiul divin, este imposibil de explicat natura omului”.
Informațiile de mai sus nu spun nimic despre asta.
Genomul, într-adevăr, are o structură mai complexă decât se credea anterior.
Dar, la urma urmei, computerul menționat în articol nu este format doar din celule de memorie.
Un computer are două memorii: pe termen lung și operațional, precum și un procesor în care sunt procesate informații. Participă la prelucrarea informațiilor și a câmpului electromagnetic. Pentru a descifra informațiile genomului, este necesar să înțelegem cum se întâmplă, nu numai stocarea informațiilor, ci și procesarea acesteia. Recunosc și ideea că o parte din informații este stocată înregistrată de câmp electromagnetic. Și, de asemenea, în afara unei persoane, așa cum am scris deja, în centrele speciale de informare ale Minții Superioare.
Imaginează-ți un text continuu codificat în cod binar 0 sau 1 în cod Morse, în timp ce nu știi în ce limbă (engleză sau franceză....) este scris și nu știi că acest text continuu este format din cuvinte , propoziții, paragrafe, capitole, volume, rafturi, dulapuri etc.
Este aproape același lucru în biologie, doar că totul este codificat aici cu un cod de patru cifre și până acum am descifrat ordinea genelor elementare + - / *, dar nu cunoaștem limba și, în consecință cuvinte, propoziții, paragrafe, capitole, volume, rafturi, dulapuri etc. Genomul descifrat pentru noi este încă un text solid al unui cod de 4 cereale și este aproape imposibil să-l studiem pe toate direct.
Dar se dovedește că în anumite momente (atât la un individ și la cohorta lui de generații, cât și la o specie, gen), unele gene și complexele acestora (responsabile de cuvinte, propoziții, paragrafe, capitole, volume, rafturi, dulapuri etc. ) sunt active, iar în alte perioade de evoluție sunt pasive, pe care le-am determinat indirect de diverse trăsături poligenice (ceea ce este arătat în subiectul Legea periodică universală a evoluției).
Până acum, există doar două metode pentru studierea genelor, acesta este un simplu calcul de laborator al sumei genelor (ADN) dintr-o probă și există un dispozitiv care numără cantitatea de ARN produsă de proteine. blocat pe cipul electronic generat ADN specific, dar din moment ce o cantitate imensă de ADN este activă în fiecare moment și, în consecință, o cantitate imensă de proteine diferite sunt produse prin ARN, este foarte dificil să se separe „acești tăiței cu o lingură, furculiță și betisoare japoneze” în această supă și găsiți ceea ce căutați foarte greu - să găsiți relații cauzale între un anumit ADN (ca complex de ADN) și influența acestuia asupra unei trăsături poligenice.
Se pare că am găsit o metodă simplă de a sorta toată această supă de ADN, ARN și proteinele lor care determină gradul unei trăsături poligenice.
După cum sa dovedit, fiecare trăsătură poligenică în ordinea evoluției unui individ (cohorte de generații, specii și gen) este periodică, prin urmare, trebuie să fie periodice în activitatea ARN și ADN și, prin urmare, este necesar doar să se găsească (în primul rând intrând în detalii genetice) o corelație între modificarea metrică într-o trăsătură poligenică (la un individ, o cohortă de generații, o specie, un gen...) și proporțională cu aceste perioade, activitatea corespunzătoare a ARN-ului, ADN-ului.
) găsit în genomul muștei fructelor ( Drosophila ananassae) o copie completă a genomului bacterian parazit Wolbachia.
Bacteria Wolbachia trăiește în citoplasma celulelor gazdă și este cunoscută pentru că a învățat să regleze fin reproducerea, dezvoltarea și chiar evoluția gazdelor sale. Prin urmare, este adesea numit „microb-manipulator” sau „stăpânul muștelor” (din moment ce trăiește în celule de insecte).
Studiul a început când Julie Dunning-Hotopp, de la JCVI, a descoperit cum anumite gene Wolbachia „cooperează” cu genele Drosophila ca și cum ar fi parte din același genom.
Michael Clark - cercetător la Universitatea din Rochester - a stabilit colonia Drosophila ananassaeîn laborator pentru a afla secretul cu Warren.
Gena Wolbachia din genomul Drosophila (ilustrat de Universitatea din Rochester).
„Timp de câteva luni, am crezut că am greșit cu ceva”, spune Clarke, „am sugerat chiar că s-a dezvoltat rezistență la antibiotice, deoarece am găsit fiecare genă Wolbachia din nou și din nou. Când am luat în sfârșit șervețele pe care le lăsasem singur cu câteva luni în urmă, nu am găsit Wolbachia în sine.
Acum Warren și Clarke încearcă să-și dea seama care este avantajul inserării unei părți atât de mari de ADN pentru Drosophila - poate că genele „străine” oferă gazdei câteva oportunități noi.

Și astfel genele Wolbachia trec în ADN-ul gazdei (ilustrare de Nicolle Rager Fuller, National Science).
Rezultatele studiului au fost publicate într-un articol din revista Science. În ea, autorii sugerează că transferul de gene orizontal (transferul de gene între specii neînrudite) are loc între bacterii și organisme multicelulare din lumea noastră mult mai des decât se credea anterior.
Descifrarea mecanismelor genetice moleculare ale manipulărilor efectuate de Wolbachia cu gazdele sale va oferi omului noi mijloace puternice de influențare a organismelor vii și a naturii în ansamblu.
Cu toate acestea, nu toate insectele sunt supuse influenței negative a Wolbachia. De exemplu, fluturii din Insulele Samoan au „învățat” să-și protejeze masculii. Mă întreb dacă țânțarii de malarie, pe care vor să-i infecteze cu această bacterie, vor învăța să lupte cu ea?
La 50 de ani de la descoperirea structurii ADN-ului
A.V. Zelenin
GENOMUL PLANTELOR
A. V. Zelenin
Zelenin Alexandru Vladimirovici- d.b.n.,
şef al laboratorului Institutului de Biologie Moleculară. V.A. Engelhardt RAS.
Realizările impresionante ale programului „Genom uman”, precum și succesul lucrărilor de descifrare a așa-numitelor genomuri extra-mici (virusuri), mici (bacterii, drojdie) și medii (viermi rotunzi, Drosophila), au făcut posibilă trecerea la un studiu pe scară largă a genomului plantelor mari și foarte mari. Necesitatea urgentă a unui studiu detaliat al genomilor celor mai importante plante din punct de vedere economic a fost subliniată la o întâlnire despre genomica plantelor, organizată în 1997 în Statele Unite [ , ]. De-a lungul anilor care au trecut de atunci, s-au obținut succese neîndoielnice în acest domeniu. În 2000, a apărut o publicație privind secvențierea completă (determinarea secvenței nucleotidice liniare a întregului ADN nuclear) a genomului muștarului mic - Arabidopsis, în 2001 - privind secvențierea preliminară (proiect) a genomului orezului. Au fost raportate în mod repetat lucrări de secvențiere a genomului plantelor mari și super-mari (porumb, secară, grâu), cu toate acestea, aceste rapoarte nu conțineau informații specifice și erau mai degrabă de natura declarațiilor de intenție.
Se presupune că decodificarea genomului plantelor va deschide perspective largi pentru știință și practică. În primul rând, identificarea de noi gene și lanțul de reglare genetică a acestora va crește semnificativ productivitatea plantelor prin utilizarea abordărilor biotehnologice. Odată cu descoperirea, izolarea, reproducerea (clonarea) și secvențierea genelor responsabile pentru funcții atât de importante ale organismului vegetal precum reproducerea și productivitatea, procesele de variabilitate, rezistența la factorii de mediu adversi, precum și împerecherea omoloagă a cromozomilor, apariția se asociază noi oportunităţi de îmbunătăţire a procesului de ameliorare . În cele din urmă, genele izolate și donate pot fi folosite pentru a obține plante transgenice cu proprietăți fundamental noi și pentru a analiza mecanismele de reglare a activității genelor.
Importanța studierii genomului plantelor este subliniată și de faptul că până în prezent numărul de gene de plante localizate, clonate și secvențiate este mic și variază, după diverse estimări, între 800 și 1200. Acesta este de 10-15 ori mai mic decât, pt. exemplu, la oameni.
Statele Unite rămân liderul incontestabil în studiul pe scară largă a genomului plantelor, deși studii intensive ale genomului orezului sunt efectuate în Japonia și, în ultimii ani, în China. La descifrarea genomului Arabidopsis, pe lângă laboratoarele din SUA, au participat activ grupurile de cercetare europene. Conducerea aparentă a Statelor Unite provoacă îngrijorare serioasă a oamenilor de știință europeni, pe care aceștia au exprimat-o în mod clar la o întâlnire sub titlul semnificativ „Perspective pentru genomica în era post-genomică”, desfășurată la sfârșitul anului 2000 în Franța. Progresul științei americane în studierea genomului plantelor agricole și crearea formelor de plante transgenice, potrivit oamenilor de știință europeni, amenință că într-un viitor nu prea îndepărtat (două-cinci decenii), când creșterea populației va pune omenirea în fața unui criza alimentară, economia și știința europeană vor deveni dependente de tehnologia americană. În acest sens, a fost anunțată crearea unui program științific franco-german pentru studiul genomului plantelor („Plantgene”) și s-au făcut investiții semnificative în acesta.
Evident, problemele genomicei plantelor ar trebui să atragă atenția oamenilor de știință și organizatorilor ruși ai științei, precum și a organelor de conducere, deoarece nu este vorba doar de prestigiul științific, ci și de securitatea națională a țării. Într-un deceniu sau două, alimentele vor deveni cea mai importantă resursă strategică.
DIFICULTĂȚI ÎN STUDIAREA GENOMULUI PLANTELOR
Studiul genomului plantelor este o sarcină mult mai dificilă decât studiul genomului oamenilor și al altor animale. Acest lucru se datorează următoarelor circumstanțe:
dimensiuni uriașe ale genomului, atingând zeci și chiar sute de miliarde de perechi de baze (pb) pentru speciile individuale de plante: genomul principalelor plante importante din punct de vedere economic (cu excepția orezului, inului și bumbacului) fie sunt apropiate ca mărime de genomul uman, fie depășește-l de multe ori (tabel);STUDII CROMOZOMIALE ALE GENOMURILORFluctuații puternice ale numărului de cromozomi la diferite plante - de la două la unele specii la câteva sute la altele și nu este posibil să se identifice o corelație strictă între dimensiunea genomului și numărul de cromozomi;
Se formează o abundență de poliploide (conținând mai mult de doi genomi per celulă) cu genomi similari, dar nu identici (alpoliploidie);
Îmbogățirea extremă a genomului plantelor (până la 99%) de ADN „nesemnificativ” (necodificare, adică neconținând gene), ceea ce face foarte dificilă unirea (aranjarea în ordinea corectă) a fragmentelor secvențiate într-un mod comun. regiune ADN de dimensiuni mari (contig);
Cartografierea morfologică, genetică și fizică incompletă a cromozomilor (comparativ cu genomul Drosophila, uman și șoarece);
Imposibilitatea practică a izolării cromozomilor individuali în formă pură folosind metode utilizate de obicei în acest scop pentru cromozomi umani și animale (sortarea în flux și utilizarea hibrizilor celulari);
Dificultatea cartografierii cromozomiale (determinarea locației pe cromozom) a genelor individuale folosind hibridizare in situ, datorită atât conținutului ridicat de ADN „nesemnificativ” din genomul plantelor, cât și particularităților organizării structurale a cromozomilor plantelor;
Depărtarea evolutivă a plantelor față de animale, ceea ce complică serios utilizarea informațiilor obținute prin secvențierea genomului oamenilor și a altor animale pentru studiul genomului plantelor;
Procesul lung de reproducere al majorității plantelor, care încetinește semnificativ analiza genetică a acestora.
Studiile cromozomiale (citogenetice) ale genomului în general și ale plantelor în special au o istorie lungă. Termenul „genom” a fost propus pentru a desemna un set haploid (unic) de cromozomi cu genele conținute în ei în primul sfert al secolului al XX-lea, adică cu mult înainte de stabilirea rolului ADN-ului ca purtător de informații genetice. .
Descrierea genomului unui organism multicelular nou, nestudiat genetic anterior, începe de obicei cu studiul și descrierea setului complet al cromozomilor săi (cariotip). Acest lucru, desigur, se aplică și plantelor, dintre care un număr mare nici măcar nu au început să fie studiate.
Deja în zorii studiilor cromozomiale, genomurile speciilor de plante înrudite au fost comparate pe baza analizei conjugării meiotice (combinație de cromozomi omologi) în hibrizi interspecifici. În ultimii 100 de ani, posibilitățile de analiză a cromozomilor s-au extins dramatic. Acum, pentru caracterizarea genomului plantelor sunt folosite tehnologii mai avansate: diverse variante ale așa-numitei colorări diferențiale, care face posibilă identificarea cromozomilor individuali după caracteristicile morfologice; hibridizare in situ făcând posibilă localizarea unor gene specifice pe cromozomi; studii biochimice ale proteinelor celulare (electroforeza si imunochimie) si, in final, un set de metode bazate pe analiza ADN-ului cromozomial pana la secventierea acestuia.

Orez. unu. Cariotipurile cerealelor a - secară (14 cromozomi), b - grâu dur (28 cromozomi), c - grâu moale (42 cromozomi), d - orz (14 cromozomi)De mulți ani, au fost studiate cariotipurile cerealelor, în primul rând grâul și secară. Interesant este că la diferite specii ale acestor plante, numărul de cromozomi este diferit, dar întotdeauna un multiplu de șapte. Tipurile individuale de cereale pot fi recunoscute în mod fiabil după cariotipul lor. De exemplu, genomul de secară este format din șapte perechi de cromozomi mari cu blocuri heterocromatice intens colorate la capete, adesea numite segmente sau benzi (Fig. 1a). Genomul grâului are deja 14 și 21 de perechi de cromozomi (Fig. 1, b, c), iar distribuția blocurilor heterocromatice în ele nu este aceeași ca și în cromozomii de secară. Genomii individuali de grâu, desemnați A, B și D, diferă, de asemenea, unul de celălalt. O creștere a numărului de cromozomi de la 14 la 21 duce la o schimbare bruscă a proprietăților grâului, care se reflectă în numele lor: durum, sau paste, grâu și moale, sau pâine, grâu . Gena D, care conține gene pentru proteinele glutenului, care conferă aluatului așa-numita germinație, este responsabilă pentru dobândirea de înalte proprietăți de coacere de către grâul moale. Acest genom este căruia i se acordă o atenție deosebită în îmbunătățirea selecției grâului de pâine. O altă cereală cu 14 cromozomi, orzul (Fig. 1, d), nu este de obicei folosită pentru a face pâine, dar este principala materie primă pentru fabricarea unor produse atât de comune precum berea și whisky-ul.
Cromozomii unor plante sălbatice utilizate pentru îmbunătățirea calității celor mai importante specii agricole, precum rudele sălbatice ale grâului - Aegilops, sunt în curs de studiu intens. Noi forme de plante sunt create prin încrucișare (Fig. 2) și selecție. În ultimii ani, o îmbunătățire semnificativă a metodelor de cercetare a făcut posibilă începerea studiului genomului plantelor, ale căror cariotipuri (în principal dimensiunea mică a cromozomilor) le făceau anterior inaccesibile pentru analiza cromozomilor. Așadar, abia recent au fost identificați pentru prima dată toți cromozomii de bumbac, mușețel și in.

Orez. 2. Cariotipuri de grâu și un hibrid de grâu cu Aegilops
a - grâu moale hexaploid ( Triticum astivum), constând din genomi A, B și O; b - grâu tetraploid ( Triticum timopheevi), constând din genomi A și G. conține gene de rezistență la majoritatea bolilor grâului; c - hibrizi Triticum astivum X Triticum timopheevi rezistent la mucegai și rugină, înlocuirea unei părți a cromozomilor este clar vizibilăSTRUCTURA PRIMARĂ A ADN-ului
Odată cu dezvoltarea geneticii moleculare, însuși conceptul de genom s-a extins. Acum acest termen este interpretat atât în sensul cromozomial clasic, cât și în sensul molecular modern: întregul material genetic al unui virus, al unei celule și al unui organism individual. Desigur, în urma studiului structurii primare complete a genomului (cum este adesea numită secvența liniară completă a bazelor de acid nucleic) a unui număr de microorganisme și oameni, a apărut problema secvențierii genomului plantelor.
Dintre numeroasele organisme vegetale, două au fost selectate pentru studiu - Arabidopsis, reprezentând clasa dicotiledoneelor (dimensiunea genomului 125 milioane bp) și orez din clasa monocotiledonelor (420-470 milioane bp). Acești genomi sunt mici în comparație cu alți genomi de plante și conțin relativ puține segmente ADN repetitive. Astfel de caracteristici au dat speranța că genomii selectați vor fi disponibili pentru determinarea relativ rapidă a structurii lor primare.

Orez. 3. Arabidopsis - muștar mic - o plantă mică din familia cruciferelor ( Brassicaceae). Pe un spațiu egal ca suprafață cu o pagină a revistei noastre, puteți crește până la o mie de organisme Arabidopsis individuale.Motivul alegerii Arabidopsis nu a fost doar dimensiunea redusă a genomului său, ci și dimensiunea redusă a organismului, ceea ce face ușoară creșterea lui în laborator (Fig. 3). Am luat în considerare ciclul său de reproducere scurt, datorită căruia este posibil să se efectueze rapid experimente privind încrucișarea și selecția, genetica studiată în detaliu, ușurința de manipulare în condițiile de creștere în schimbare (schimbarea compoziției de sare a solului, adăugarea diverșilor nutrienți etc. .) și testarea efectului asupra plantelor a diverșilor factori mutageni și agenți patogeni (viruși, bacterii, ciuperci). Arabidopsis nu are valoare economică, prin urmare, genomul său, împreună cu genomul șoarecelui, a fost numit referință sau, mai puțin precis, model.*
* Apariția termenului „genom model” în literatura rusă este rezultatul unei traduceri inexacte a expresiei engleze model genom. Cuvântul „model” înseamnă nu numai adjectivul „model”, ci și substantivul „probă”, „standard”, „model”. Ar fi mai corect să vorbim despre un genom eșantion sau despre un genom de referință.Lucrările intensive privind secvențierea genomului Arabidopsis au fost începute în 1996 de un consorțiu internațional care includea instituții științifice și grupuri de cercetare din SUA, Japonia, Belgia, Italia, Marea Britanie și Germania. În decembrie 2000, au devenit disponibile informații extinse care rezumă determinarea structurii primare a genomului Arabidopsis. Pentru secvențiere a fost folosită tehnologia clasică sau ierarhică: mai întâi, au fost studiate secțiuni individuale mici ale genomului, din care au fost compuse secțiuni mai mari (contigs) și, în stadiul final, structura cromozomilor individuali. ADN-ul nuclear al genomului Arabidopsis este distribuit pe cinci cromozomi. În 1999, au fost publicate rezultatele secvențierii a doi cromozomi, iar apariția în presă a informațiilor despre structura primară a celor trei rămași a completat secvențierea întregului genom.
Din 125 de milioane de perechi de baze, a fost determinată structura primară de 119 milioane, ceea ce reprezintă 92% din întregul genom. Doar 8% din genomul Arabidopsis care conține blocuri mari de segmente repetitive de ADN s-a dovedit a fi inaccesibil pentru studiu. În ceea ce privește completitatea și minuțiozitatea secvențierii genomului eucariotic, Arabidopsis rămâne în primii trei campioni împreună cu un organism de drojdie unicelulară. Saccharomyces cerevisiaeși organism multicelular Caenorhabditis eleganta(Vezi tabelul).
Aproximativ 15.000 de gene individuale care codifică proteine au fost găsite în genomul Arabidopsis. Aproximativ 12 000 dintre acestea sunt conținute sub formă de două copii per genom haploid (unic), astfel încât numărul total de gene este de 27 000. Numărul de gene din Arabidopsis nu diferă mult de numărul de gene din organisme precum oamenii și șoarecii, dar dimensiunea genomului său de 25-30 de ori mai mică. Această circumstanță este asociată cu caracteristici importante în structura genelor individuale Arabidopsis și structura generală a genomului său.
Genele Arabidopsis sunt compacte, conținând doar câțiva exoni (regiuni care codifică proteine) separați de segmente scurte (aproximativ 250 bp) de ADN necodificatori (introni). Intervalele dintre genele individuale sunt în medie de 4600 de perechi de baze. Pentru comparație, subliniem că genele umane conțin multe zeci și chiar sute de exoni și introni, iar regiunile intergenice au dimensiuni de 10 mii de perechi de baze sau mai mult. Se presupune că prezența unui genom compact mic a contribuit la stabilitatea evolutivă a Arabidopsis, deoarece ADN-ul său a devenit o țintă pentru diferiți agenți dăunători într-o măsură mai mică, în special pentru introducerea de fragmente de ADN repetitive asemănătoare virusului (transpozoni). în genom.
Printre alte caracteristici moleculare ale genomului Arabidopsis, trebuie remarcat faptul că exonii sunt îmbogățiți în guanină și citozină (44% în exoni și 32% în introni) în comparație cu genele animale, precum și prezența genelor dublu repetate (duplicate). Se presupune că o astfel de dublare a avut loc ca urmare a a patru evenimente simultane, constând în dublarea (repetarea) unei părți a genelor Arabidopsis sau fuziunea genomilor înrudite. Aceste evenimente, care au avut loc în urmă cu 100-200 de milioane de ani, sunt o manifestare a tendinței generale de poliploidizare (o creștere multiplă a numărului de genomi dintr-un organism), care este caracteristică genomului plantelor. Cu toate acestea, unele fapte arată că genele duplicate în Arabidopsis nu sunt identice și funcționează diferit, ceea ce poate fi asociat cu mutații în regiunile lor de reglementare.
Orezul a devenit un alt obiect de secvențiere completă a ADN-ului. Genomul acestei plante este, de asemenea, mic (12 cromozomi, dând un total de 420-470 milioane bp), doar de 3,5 ori mai mare decât cel al Arabidopsis. Cu toate acestea, spre deosebire de Arabidopsis, orezul are o mare importanță economică, fiind baza de nutriție pentru mai mult de jumătate din umanitate, prin urmare, nu numai miliarde de consumatori, ci și o armată de oameni multimilionare implicați activ în procesul foarte laborios al acestuia. cultivarea sunt extrem de interesați să-și îmbunătățească proprietățile.
Unii cercetători au început să studieze genomul orezului încă din anii 1980, dar aceste studii au atins o amploare serioasă abia în anii 1990. În 1991, în Japonia a fost creat un program de descifrare a structurii genomului orezului, reunind eforturile multor grupuri de cercetare. În 1997, pe baza acestui program a fost organizat Proiectul Internațional al Genomului Orezului. Participanții săi au decis să-și concentreze eforturile pe secvențierea uneia dintre subspeciile de orez ( Oriza sativajaponica), în studiul căruia s-au înregistrat deja progrese semnificative până la acel moment. Un stimulent serios și, la figurat vorbind, o vedetă călăuzitoare pentru o astfel de muncă a fost programul „Genom uman”.
În cadrul acestui program a fost testată strategia de diviziune ierarhică „cromozomială” a genomului, pe care participanții consorțiului internațional au folosit-o pentru a descifra genomul orezului. Cu toate acestea, dacă în studiul genomului uman, fracțiile de cromozomi individuali au fost izolate folosind diverse metode, atunci materialul specific pentru cromozomii individuali de orez și regiunile lor individuale a fost obținut prin microdisecție cu laser (decuparea obiectelor microscopice). Pe o lamă de microscop, unde se află cromozomii de orez, sub influența unui fascicul laser, totul este ars, cu excepția cromozomului sau a secțiunilor acestuia programate pentru analiză. Materialul rămas este folosit pentru clonare și secvențiere.
Au fost publicate numeroase rapoarte cu privire la rezultatele secvențierii fragmentelor individuale ale genomului orezului, efectuate cu mare acuratețe și detaliu, caracteristice tehnologiei ierarhice. Se credea că determinarea structurii primare complete a genomului orezului va fi finalizată până la sfârșitul anului 2003 – mijlocul anului 2004, iar rezultatele, împreună cu datele privind structura primară a genomului Arabidopsis, vor fi utilizate pe scară largă în comparație. genomica altor plante.
Cu toate acestea, la începutul anului 2002, două grupuri de cercetare - unul din China, celălalt din Elveția și Statele Unite - au publicat rezultatele unui proiect complet de secvențiere (aproximativă) a genomului orezului, realizat folosind tehnologia de clonare totală. Spre deosebire de studiul în etape (ierarhice), abordarea totală se bazează pe clonarea simultană a întregului ADN genomic într-unul dintre vectorii virali sau bacterieni și obținerea unui număr semnificativ (uriaș pentru genomi medii și mari) de clone individuale care conțin diverse Segmente de ADN. Pe baza analizei acestor secțiuni secvențiate și a suprapunerii secțiunilor terminale identice ale ADN-ului, se formează un contig - un lanț de secvențe ADN unite între ele. Contigul general (total) este structura primară a întregului genom sau cel puțin a unui cromozom individual.
Într-o astfel de prezentare schematică, strategia clonării totale pare simplă. De fapt, întâmpină dificultăți serioase asociate cu necesitatea obținerii unui număr imens de clone (se acceptă în general că genomul sau regiunea sa studiată trebuie să fie suprapuse de clone de cel puțin 10 ori), o cantitate imensă de secvențiere și extrem de complexă. munca la clone de andocare care necesită participarea specialiștilor în bioinformatică. Un obstacol serios în calea clonării totale este o varietate de segmente ADN repetitive, al căror număr, după cum sa menționat deja, crește brusc pe măsură ce dimensiunea genomului crește. Prin urmare, strategia de secvențiere totală este utilizată în principal în studiul genomului virusurilor și microorganismelor, deși a fost folosită cu succes pentru a studia genomul unui organism multicelular, Drosophila.
Rezultatele secvențierii totale a acestui genom au fost „suprapuse” unei game uriașe de informații despre structura sa cromozomială, genică și moleculară, obținute pe o perioadă de aproape 100 de ani de studiu a Drosophila. Și totuși, în ceea ce privește gradul de secvențiere, genomul Drosophila (66% din dimensiunea totală a genomului) este semnificativ inferior genomul Arabidopsis (92%), în ciuda dimensiunilor lor destul de apropiate - 180 de milioane și, respectiv, 125 de milioane de perechi de baze. . Prin urmare, recent s-a propus denumirea tehnologiei mixte, care a fost folosită pentru secvențierea genomului Drosophila.
Pentru a secvenționa genomul orezului, grupurile de cercetare menționate mai sus au luat două dintre subspeciile sale, cele mai cultivate în țările asiatice, - Oriza saliva L. ssp indicajși Oriza saliva L. sspjaponica. Rezultatele studiilor lor coincid în multe privințe, dar diferă în multe privințe. Astfel, reprezentanții ambelor grupuri au declarat că au ajuns la aproximativ 92-93% din suprapunerea genomului cu contigs. S-a demonstrat că aproximativ 42% din genomul orezului este reprezentat de repetări scurte de ADN constând din 20 de perechi de baze, iar majoritatea elementelor mobile de ADN (transpozoni) sunt localizate în regiuni intergenice. Cu toate acestea, datele privind dimensiunea genomului orezului diferă semnificativ.
Pentru subspecia japoneză, dimensiunea genomului este determinată a fi de 466 milioane de perechi de baze, iar pentru subspecia indiană, 420 milioane. Motivul acestei discrepanțe nu este clar. Poate fi o consecință a diferitelor abordări metodologice în determinarea dimensiunii părții necodificatoare a genomului, adică nu reflectă adevărata stare a lucrurilor. Dar este posibil să existe o diferență de 15% în dimensiunea genomilor studiati.
A doua discrepanță majoră a fost dezvăluită în numărul de gene găsite: pentru subspecia japoneză, de la 46 022 la 55 615 gene per genom, iar pentru subspecia indiană, de la 32 000 la 50 000. Motivul acestei discrepanțe nu este clar.
Incompletitudinea și inconsecvența informațiilor primite se remarcă în comentariile la articolele publicate. De asemenea, aici se exprimă speranța că lacunele în cunoașterea genomului orezului vor fi eliminate prin compararea datelor de „secvențiere brută” cu rezultatele secvențierii detaliate, ierarhice, efectuate de participanții la Proiectul Internațional de Genom al Orezului.
GENOMIA PLANTELOR COMPARATIVA SI FUNCTIONALA
Datele extinse obținute, dintre care jumătate (rezultatele grupului chinez) sunt disponibile publicului, deschid fără îndoială perspective largi atât pentru studiul genomului orezului, cât și pentru genomica plantelor în general. O comparație a proprietăților genomului Arabidopsis și al orezului a arătat că majoritatea genelor (până la 80%) identificate în genomul Arabidopsis se găsesc și în genomul orezului, totuși, pentru aproximativ jumătate din genele găsite în orez, analogi (ortologi) ) nu au fost încă găsite în genomul Arabidopsis. În același timp, 98% dintre genele a căror structură primară a fost stabilită pentru alte cereale au fost găsite în genomul orezului.
Discrepanța semnificativă (aproape dublă) dintre numărul de gene din orez și Arabidopsis este uluitoare. În același timp, datele proiectului de decodificare a genomului orezului, obținute prin secvențierea totală, practic nu sunt comparate cu rezultatele ample ale studiului genomului orezului prin metoda clonării și secvențierii ierarhice, adică ceea ce are nu a fost efectuată cu privire la genomul Drosophila. Prin urmare, rămâne neclar dacă diferența în numărul de gene în Arabidopsis și orez reflectă adevărata stare de lucruri sau dacă este explicată prin diferența de abordări metodologice.
Spre deosebire de genomul Arabidopsis, nu sunt date date despre genele gemene din genomul orezului. Este posibil ca cantitatea lor relativă să fie mai mare în orez decât în Arabidopsis. Această posibilitate este susținută indirect de date privind prezența formelor poliploide de orez. Se poate aștepta mai multă claritate cu privire la această problemă după finalizarea Proiectului Internațional de Genom al Orezului și se obține o imagine detaliată a structurii primare a ADN-ului acestui genom. Motive serioase pentru o astfel de speranță sunt oferite de faptul că, după publicarea lucrărilor privind secvențierea brută a genomului de orez, numărul de publicații privind structura acestui genom a crescut brusc, în special, au apărut informații despre secvențierea detaliată. dintre cromozomii 1 și 4 ai săi.
Cunoașterea, cel puțin aproximativ, a numărului de gene din plante este de o importanță fundamentală pentru genomica comparativă a plantelor. Inițial, s-a crezut că, deoarece toate plantele cu flori sunt foarte apropiate una de cealaltă în ceea ce privește caracteristicile lor fenotipice, genomul lor ar trebui să fie, de asemenea, similar. Și dacă studiem genomul Arabidopsis, vom obține informații despre majoritatea genomilor altor plante. O confirmare indirectă a acestei presupuneri sunt rezultatele secvențierii genomului șoarecelui, care este surprinzător de aproape de genomul uman (aproximativ 30 de mii de gene, dintre care doar 1 mie s-au dovedit a fi diferite).
Se poate presupune că motivul diferențelor dintre genomul Arabidopsis și orezul constă în apartenența lor la diferite clase de plante - dicotiledone și monocotiledone. Pentru a clarifica această problemă, este foarte de dorit să se cunoască cel puțin o structură primară brută a unei alte plante monocotiledonate. Cel mai realist candidat ar putea fi porumbul, al cărui genom este aproximativ egal cu genomul uman, dar totuși mult mai mic decât genomul altor cereale. Valoarea nutritivă a porumbului este binecunoscută.
Vastul material obținut ca urmare a secvențialării genomului Arabidopsis și orezului devine treptat baza unui studiu la scară largă al genomului plantelor folosind genomica comparativă. Astfel de studii au o semnificație biologică generală, deoarece fac posibilă stabilirea principiilor principale ale organizării genomului plantei în ansamblu și a cromozomilor lor individuali, identificarea trăsăturilor comune ale structurii genelor și regiunilor lor de reglare și luarea în considerare a acestora. raportul dintre partea activă funcțional (genă) a cromozomului și diferitele regiuni ADN intergenice care nu codifică proteine. Genetica comparativă devine, de asemenea, din ce în ce mai importantă pentru dezvoltarea genomicii funcționale umane. Pentru studii comparative s-a efectuat secvențierea genomilor de pește-papă și șoarece.
La fel de important este studiul genelor individuale responsabile de sinteza proteinelor individuale care determină funcțiile specifice ale corpului. Este în descoperirea, izolarea, secvențierea și determinarea funcției genelor individuale în care se află semnificația practică, în primul rând medicală, a programului Genomului uman. Această împrejurare a fost remarcată în urmă cu câțiva ani de către J. Watson, care a subliniat că programul „Genom uman” va fi finalizat numai atunci când funcțiile tuturor genelor umane vor fi determinate.
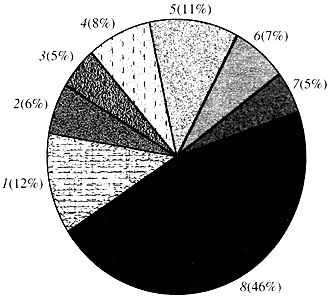
Orez. patru. Clasificare în funcție de funcția genelor Arabidopsis
1 - gene pentru creștere, diviziune și sinteza ADN; 2 - gene de sinteză a ARN (transcripție); 3 - gene pentru sinteza si modificarea proteinelor; 4 - gene pentru dezvoltare, îmbătrânire și moarte celulară; 5 - gene ale metabolismului celular și ale metabolismului energetic; 6 - gene de interacțiune intercelulară și transmitere a semnalului; 7 - gene pentru furnizarea altor procese celulare; 8 - gene cu funcție necunoscutăÎn ceea ce privește funcția genelor plantelor, știm mai puțin de o zecime din ceea ce știm despre genele umane. Chiar și în Arabidopsis, al cărui genom este mult mai studiat decât genomul uman, funcția a aproape jumătate din genele sale rămâne necunoscută (Fig. 4). Între timp, pe lângă genele comune animalelor, plantele au un număr semnificativ de gene care sunt specifice doar (sau cel puțin predominant) pentru ele. Vorbim despre genele implicate în transportul apei și sinteza peretelui celular, care este absent la animale, despre genele care asigură formarea și funcționarea cloroplastelor, fotosinteza, fixarea azotului, precum și sinteza a numeroase produse aromatice. Această listă poate fi continuată, dar este deja clar cu ce sarcină dificilă se confruntă genomica funcțională a plantelor.
Secvențierea întregului genom oferă informații aproape adevărate despre numărul total de gene dintr-un organism dat, face posibilă plasarea unor informații mai mult sau mai puțin detaliate și fiabile despre structura lor în băncile de date și facilitează munca de izolare și studiere a genelor individuale. Cu toate acestea, secvențierea genomului nu înseamnă în niciun caz să stabilească funcția tuturor genelor.
Una dintre cele mai promițătoare abordări ale genomicii funcționale se bazează pe identificarea genelor de lucru care sunt utilizate pentru transcrierea (citirea) ARNm. Această abordare, inclusiv utilizarea tehnologiei moderne de microarray, face posibilă identificarea simultană a până la zeci de mii de gene funcționale. Recent, folosind această abordare, a început studiul genomului plantelor. Pentru Arabidopsis, a fost posibil să se obțină aproximativ 26 de mii de transcrieri individuale, ceea ce facilitează foarte mult posibilitatea de a determina funcția aproape tuturor genelor sale. La cartofi, a fost posibil să se identifice aproximativ 20.000 de gene de lucru care sunt importante pentru înțelegerea atât a proceselor de creștere și formare a tuberculilor, cât și a proceselor de boală a cartofului. Se presupune că aceste cunoștințe vor crește rezistența unuia dintre cele mai importante produse alimentare la agenți patogeni.
Dezvoltarea logică a genomicii funcționale a fost proteomica. Acest nou domeniu de știință studiază proteomul, care este de obicei înțeles ca un set complet de proteine dintr-o celulă la un anumit moment. Un astfel de set de proteine, care reflectă starea funcțională a genomului, se schimbă tot timpul, în timp ce genomul rămâne neschimbat.
Studiul proteinelor a fost folosit de mult timp pentru a judeca activitatea genomului plantelor. După cum se știe, enzimele prezente în toate plantele diferă în fiecare specie și soiuri în secvența de aminoacizi. Astfel de enzime, cu aceeași funcție, dar cu o secvență diferită de aminoacizi individuali, se numesc izoenzime. Au proprietăți fizico-chimice și imunologice diferite (greutate moleculară, sarcină), care pot fi detectate prin cromatografie sau electroforeză. De mulți ani, aceste metode au fost folosite cu succes pentru a studia așa-numitul polimorfism genetic, adică diferențele dintre organisme, soiuri, populații, specii, în special grâul și formele înrudite de cereale. Recent, însă, datorită dezvoltării rapide a metodelor de analiză a ADN-ului, inclusiv secvențierea, studiul polimorfismului proteic a fost înlocuit cu studiul polimorfismului ADN. Cu toate acestea, studiul direct al spectrelor proteinelor de depozitare (prolamine, gliadine etc.), care determină principalele proprietăți nutriționale ale cerealelor, rămâne o metodă importantă și de încredere pentru analiza genetică, selecția și producția de semințe a plantelor agricole.
Cunoașterea genelor, a mecanismelor de exprimare și reglare a acestora este extrem de importantă pentru dezvoltarea biotehnologiei și producerea de plante transgenice. Se știe că succesele impresionante în acest domeniu provoacă o reacție ambiguă din partea comunității de mediu și medicale. Cu toate acestea, există o zonă a biotehnologiei vegetale în care aceste temeri, dacă nu complet nefondate, atunci, în orice caz, par a fi de puțină importanță. Vorbim despre crearea de plante industriale transgenice care nu sunt folosite ca produse alimentare. India a recoltat recent prima recoltă de bumbac transgenic care este rezistent la o serie de boli. Există informații despre introducerea unor gene speciale care codifică proteine pigmentare în genomul bumbacului și producerea de fibre de bumbac care nu necesită vopsire artificială. O altă cultură industrială care poate face obiectul unei inginerie genetică eficientă este inul. Utilizarea sa ca alternativă la bumbac pentru materii prime textile a fost discutată recent. Această problemă este extrem de importantă pentru țara noastră, care și-a pierdut propriile surse de bumbac brut.
PERSPECTIVE DE STUDIAREA GENOMULUI PLANTELOR
Evident, studiile structurale ale genomului plantelor se vor baza pe abordările și metodele genomicei comparative, folosind ca material principal rezultatele descifrării genomului Arabidopsis și orezului. Un rol important în dezvoltarea genomicii comparative a plantelor îl vor juca, fără îndoială, informațiile care mai devreme sau mai târziu vor fi furnizate de secvențierea totală (aproximată) a genomilor altor plante. În acest caz, genomica comparativă a plantelor se va baza pe stabilirea de relații genetice între loci individuali și cromozomi aparținând unor genomi diferiți. Ne vom concentra nu atât pe genomica generală a plantelor, cât pe genomica selectivă a locurilor cromozomiale individuale. De exemplu, s-a demonstrat recent că gena responsabilă pentru vernalizare este localizată la locusul VRn-AI al cromozomului hexaploid de grâu 5A și locusul Hd-6 al cromozomului 3 de orez.
Dezvoltarea acestor studii va fi un impuls puternic pentru identificarea, izolarea și secvențierea multor gene ale plantelor importante din punct de vedere funcțional, în special, genele responsabile de rezistența la boli, rezistența la secetă și adaptabilitatea la diferite condiții de creștere. Din ce în ce mai mult, se va folosi genomica funcțională, bazată pe detectarea în masă (screening) a genelor care funcționează în plante.
Putem prevedea îmbunătățiri suplimentare ale tehnologiilor cromozomiale, în primul rând metoda de microdisecție. Utilizarea sa extinde dramatic posibilitățile cercetării genomice fără a necesita costuri uriașe, cum ar fi, de exemplu, secvențierea totală a genomului. Metoda de localizare pe cromozomii plantelor a genelor individuale cu ajutorul hibridizării va fi răspândită în continuare. in situ.În prezent, utilizarea sa este limitată de numărul mare de secvențe repetitive din genomul plantei și, eventual, de particularitățile organizării structurale a cromozomilor plantelor.
Tehnologiile cromozomiale vor deveni de mare importanță pentru genomica evolutivă a plantelor în viitorul apropiat. Aceste tehnologii relativ ieftine fac posibilă evaluarea rapidă a variabilității intra și interspecifice, studiul genomilor alopoliploizi complexe de grâu tetraploid și hexaploid, triticale; analiza proceselor evolutive la nivel cromozomial; investighează formarea genomilor sintetici și introducerea (introgresiunea) materialului genetic străin; identificarea relațiilor genetice dintre cromozomii individuali ai diferitelor specii.
Studiul cariotipului plantelor folosind metode citogenetice clasice, îmbogățit prin analiză biologică moleculară și tehnologie computerizată, va fi utilizat pentru caracterizarea genomului. Acest lucru este deosebit de important pentru studierea stabilității și variabilității cariotipului la nivelul nu numai al organismelor individuale, ci și al populațiilor, soiurilor și speciilor. În cele din urmă, este dificil de imaginat cum poate fi estimat numărul și spectrele de rearanjamente cromozomiale (aberații, punți) fără utilizarea metodelor de colorare diferențială. Astfel de studii sunt extrem de promițătoare pentru monitorizarea mediului în funcție de starea genomului plantei.
În Rusia modernă, secvențierea directă a genomului plantelor este puțin probabil să fie efectuată. O astfel de muncă, care necesită investiții mari, depășește puterea economiei noastre actuale. Între timp, datele privind structura genomului Arabidopsis și orezului, obținute de știința mondială și disponibile în băncile internaționale de date, sunt suficiente pentru dezvoltarea genomicii plantelor interne. Se poate prevedea extinderea studiilor asupra genomului plantelor bazate pe abordări genomice comparative pentru a rezolva probleme specifice de ameliorare și producție a culturilor, precum și pentru a studia originea diferitelor specii de plante de mare importanță economică.
Se poate presupune că abordările genomice precum tiparea genetică (analize RELF, RAPD, AFLP etc.), care sunt destul de accesibile pentru bugetul nostru, vor fi utilizate pe scară largă în practica internă de ameliorare și producția de culturi. În paralel cu metodele directe de determinare a polimorfismului ADN, abordări bazate pe studiul polimorfismului proteic, în primul rând proteinele de depozitare a cerealelor, vor fi utilizate pentru rezolvarea problemelor de genetică și ameliorare a plantelor. Tehnologiile cromozomiale vor fi utilizate pe scară largă. Sunt relativ ieftine, dezvoltarea lor necesită investiții destul de moderate. În domeniul studiilor cromozomiale, știința domestică nu este inferioară lumii.
Trebuie subliniat faptul că știința noastră a adus o contribuție semnificativă la formarea și dezvoltarea genomicii plantelor [ , ].
Rolul fundamental l-a jucat N.I. Vavilov (1887-1943).
În biologia moleculară și genomica plantelor, contribuția de pionierat a lui A.N. Belozerski (1905-1972).
În domeniul studiilor cromozomiale, este de remarcat munca remarcabilului genetician S.G. Navashin (1857-1930), care a descoperit primul cromozomi sateliti la plante și a demonstrat că este posibil să se facă distincția între cromozomi individuali în funcție de caracteristicile morfologiei lor.
Un alt clasic al științei ruse G.A. Levitsky (1878-1942) a descris în detaliu cromozomii de secară, grâu, orz, mazăre și sfeclă de zahăr, a introdus termenul „cariotip” în știință și a dezvoltat doctrina acestuia.
Specialiștii moderni, bazându-se pe realizările științei mondiale, pot aduce o contribuție semnificativă la dezvoltarea ulterioară a geneticii și genomicii plantelor.
Autorul își exprimă sincere mulțumiri academicianului Yu.P. Altukhov pentru discuția critică a articolului și sfaturi valoroase.Activitatea echipei conduse de autorul articolului a fost susținută de Fundația Rusă pentru Cercetare de bază (granturi nr. 99-04-48832; 00-04-49036; 00-04-81086), Programul președintelui Federația Rusă pentru a sprijini școlile științifice (granturile nr. 00-115 -97833 și NSh-1794.2003.4) și Programul Academiei Ruse de Științe „Markeri genetici moleculari și cromozomiali în dezvoltarea metodelor moderne de reproducere și producție de semințe” .
LITERATURĂ
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Introducere în genomica plantelor // Biologie moleculară. 2001. V. 35. S. 339-348. 2. Pen E. Bonanza pentru genomica plantelor // Știință. 1998. V. 282. P. 652-654. 3. Genomica plantelor, Proc. Natl. Acad. sci. STATELE UNITE ALE AMERICII. 1998. V. 95. P. 1962-2032. 4. Cartel N.A. si etc. Genetica. Dicţionar enciclopedic. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferențierea genomului în Aegilops. 1. Distribuția secvențelor de ADN foarte repetitive pe cromozomii speciilor diploide, Genomul. 1996. V. 39. P. 293-306. Istoricul analizei cromozomilor // Biol. membranelor. 2001. T. 18. S. 164-172.
Gene saritoare
La mijlocul secolului trecut, cercetătorul american Barbara McClintock a descoperit gene uimitoare în porumb care își pot schimba în mod independent poziția pe cromozomi. Acum sunt numite „gene de săritură” sau elemente transpozabile (mobile). Descoperirea nu a fost recunoscută de mult timp, considerând elementele mobile un fenomen unic, caracteristic doar porumbului. Cu toate acestea, tocmai pentru această descoperire din 1983 a fost premiat McClintock Premiul Nobel Astăzi, genele de săritură au fost găsite la aproape toate speciile de animale și plante studiate.
De unde provin genele hopuri, ce fac ele în celulă, există beneficii din ele? De ce, cu părinți sănătoși din punct de vedere genetic, familia muștei de fructe Drosophila, datorită genelor săritoare, poate produce descendenți mutanți cu o frecvență ridicată sau chiar să fie complet fără copii? Care este rolul genelor săritoare în evoluție?
Trebuie spus că genele care asigură funcționarea celulelor sunt localizate pe cromozomi într-o anumită ordine. Datorită acestui fapt, pentru multe tipuri de organisme unicelulare și multicelulare, a fost posibilă construirea așa-numitelor hărți genetice. Cu toate acestea, există un ordin de mărime mai mult material genetic între gene decât în sine! Ce rol joacă această parte „balast” a ADN-ului nu a fost pe deplin stabilit, dar aici se găsesc cel mai des elementele mobile, care nu numai că se mișcă singure, ci pot lua cu ele și fragmente de ADN vecine.

De unde provin genele jumper? Se crede că cel puțin unele dintre ele sunt derivate din viruși, deoarece unele elemente mobile sunt capabile să formeze particule virale (de exemplu, elementul mobil țigan din musca fructelor). Drosophila melanogaster). Unele elemente transpozabile apar în genom prin așa-numitele transfer orizontal din alte tipuri. De exemplu, se constată că mobil vagabond-element (tradus în rusă, se numește vagabond) Drosophila melanogaster introdus în mod repetat în genomul acestei specii. Există o versiune conform căreia unele regiuni de reglementare ale ADN-ului pot avea, de asemenea, autonomie și o tendință de „vagabondaj”.
balast util
Pe de altă parte, majoritatea genelor de săritură, în ciuda numelui, se comportă liniștit, deși reprezintă o cincime din materialul genetic total. Drosophila melanogaster sau aproape jumătate din genomul uman.
Redundanța ADN-ului, care a fost menționată mai sus, are propriul plus: ADN-ul de balast (inclusiv elementele mobile pasive) ia lovitura dacă ADN străin este introdus în genom. Probabilitatea ca un nou element să fie inserat într-o genă utilă și astfel să-i perturbe funcționarea este redusă dacă există ADN mult mai voluminos decât semnificativ.
O anumită redundanță a ADN-ului este utilă în același mod ca „redundanța” literelor în cuvinte: scriem „Maria Ivanovna” și spunem „Marivana”. Unele dintre litere se pierd inevitabil, dar sensul rămâne. Același principiu funcționează și la nivelul semnificației aminoacizilor individuali dintr-o moleculă de proteină-enzimă: doar secvența de aminoacizi care formează centrul activ este strict conservatoare. Astfel, la diferite niveluri, redundanța se dovedește a fi un fel de buffer care oferă o marjă de siguranță pentru sistem. Așa se face că elementele mobile care și-au pierdut mobilitatea nu sunt inutile pentru genom. După cum se spune, „de la o oaie subțire chiar și un smoc de lână”, deși, poate, un alt proverb ar fi mai potrivit aici - „fiecare bast într-o linie”.

Elementele mobile care și-au păstrat capacitatea de a sări se mișcă de-a lungul cromozomilor Drosophila la o frecvență de 10–2–10–5 per genă pe generație, în funcție de tipul de element, fundalul genetic și conditii externe. Aceasta înseamnă că una din o sută de gene săritoare dintr-o celulă își poate schimba poziția după următoarea diviziune celulară. Ca urmare, după câteva generații, distribuția elementelor transpozabile de-a lungul cromozomului se poate schimba foarte semnificativ.
Este convenabil să se studieze o astfel de distribuție pe cromozomi politenici (multifilamentoși) din glandele salivare ale larvelor de Drosophila. Acești cromozomi sunt de multe ori mai groși decât cei normali, ceea ce face mult mai ușor de examinat la microscop. Cum sunt formați acești cromozomi? În celulele glandelor salivare, ADN-ul fiecărui cromozom este înmulțit, ca în diviziunea celulară normală, dar celula în sine nu se împarte. Ca urmare, numărul de celule din glandă nu se modifică, dar în 10-11 cicluri, câteva mii de fire de ADN identice se acumulează în fiecare cromozom.

Parțial din cauza cromozomilor politenici, genele de săritură din Drosophila sunt mai bine înțelese decât în alte metazoare. În urma acestor studii, s-a dovedit că chiar și în cadrul aceleiași populații de Drosophila este dificil să găsești doi indivizi care au cromozomi cu aceeași distribuție a elementelor mobile. Nu este o coincidență că se crede că majoritatea mutațiilor spontane la Drosophila sunt cauzate de mișcarea acestor „pâlnii”.
Consecințele pot varia...
Pe baza efectului lor asupra genomului, elementele active transpozabile pot fi împărțite în mai multe grupuri. Unele dintre ele îndeplinesc funcții extrem de importante și utile pentru genom. De exemplu, telomeric ADN-ul, situat la capetele cromozomilor, în Drosophila este format doar din elemente mobile speciale. Acest ADN este extrem de important - pierderea acestuia atrage după sine pierderea întregului cromozom în procesul de diviziune celulară, ceea ce duce la moartea celulei.
Alte elemente mobile sunt de-a dreptul „dăunători”. Cel puțin sunt considerați ca atare acest moment. De exemplu, elementele transpozabile ale clasei R2 pot fi introduse în mod specific în genele artropodelor care codifică una dintre proteinele ribozomilor - „fabrici” celulare pentru sinteza proteinelor. Indivizii cu astfel de tulburări supraviețuiesc doar pentru că doar o parte din numeroasele gene care codifică aceste proteine este deteriorată în genom.

Există și astfel de elemente mobile care se mișcă doar în țesuturile reproductive care produc celule germinale. Acest lucru se explică prin faptul că în țesuturi diferite același element mobil poate produce lungimi și funcții diferite ale moleculei proteine-enzime necesare mișcării.
Un exemplu al acestuia din urmă este elementul P Drosophila melanogaster, care a intrat în populațiile sale naturale prin transfer orizontal de la o altă specie de Drosophila cu nu mai mult de o sută de ani în urmă. Cu toate acestea, nu există aproape o populație pe Pământ în acest moment. Drosophila melanogaster, în care nu ar exista un element P. În același timp, trebuie remarcat faptul că majoritatea copiilor sale sunt defecte, în plus, aceeași versiune a defectului a fost găsită aproape peste tot. Rolul acestuia din urmă în genom este deosebit: este „intolerantă” cu semenii săi și joacă rolul unui represor, blocându-le mișcarea. Așadar, protecția genomului Drosophila de săriturile „extratereștrilor” poate fi realizată parțial prin propriii derivați.
Principalul lucru este să alegi părinții potriviți!
Majoritatea salturilor elementelor mobile nu afectează aspect Drosophila, pentru că cade pe ADN-ul de balast, dar sunt și alte situații când activitatea lor crește dramatic.

Destul de ciudat, cel mai puternic factor care induce mișcarea genelor de săritură este parentingul slab. De exemplu, ce se întâmplă dacă încrucișați femele dintr-o populație de laborator Drosophila melanogaster, care nu au un element P (pentru că strămoșii lor au fost prinși din natură în urmă cu aproximativ o sută de ani), cu masculi purtând un element P? La hibrizi, datorită mișcării rapide a elementului mobil, pot apărea un număr mare de diverse tulburări genetice. Acest fenomen, numit disgeneză hibridă, este cauzat de absența unui represor în citoplasma maternă care interzice mișcarea elementului mobil.
Astfel, dacă mirii din populația A și miresele din populația B pot crea familii numeroase, atunci inversul nu este întotdeauna adevărat. O familie de părinți sănătoși din punct de vedere genetic poate produce un număr mare de descendenți mutanți sau infertili sau chiar poate fi fără copii dacă tatăl și mama au un set diferit de elemente mobile în genom. Mai ales multe încălcări apar dacă experimentul este efectuat la o temperatură de 29 ° C. Influența factorilor externi, suprapusi pe fundalul genetic, sporește efectul nepotrivirii genomului, deși acești factori singuri (chiar și radiațiile ionizante) singuri nu sunt capabili. de a provoca o astfel de mișcare masivă a elementelor mobile.
Evenimente similare în Drosophila melanogaster poate apărea cu participarea altor familii de elemente mobile.
Evoluție „mobil”.
Genomul celular poate fi privit ca un fel de ecosistem de membri permanenți și temporari, în care vecinii nu numai că coexistă, ci și interacționează între ei. Interacțiunea genelor gazdă cu elementele transpozabile este încă puțin înțeleasă, dar pot fi citate multe rezultate - de la moartea unui organism în caz de deteriorare a unei gene importante până la refacerea funcțiilor deteriorate anterior.
Se întâmplă ca genele sărituri în sine interacționeze între ele. Astfel, se cunoaște un fenomen asemănător imunității, atunci când un element mobil nu poate fi introdus în imediata apropiere a unuia existent. Cu toate acestea, nu toate elementele mobile sunt atât de delicate: de exemplu, elementele P se pot încorpora cu ușurință unele în altele și își pot scoate frații din joc.

În plus, există un fel de autoreglare a numărului de elemente transpozabile din genom. Faptul este că elementele mobile pot schimba regiuni omoloage între ele - acest proces se numește recombinare. Ca urmare a unei astfel de interacțiuni, elementele mobile pot, în funcție de orientarea lor, să piardă ( ştergere) sau extinde ( inversiune) fragmente de ADN gazdă situate între ele. Dacă se pierde o parte semnificativă a cromozomului, genomul va muri. In cazul unei inversiuni sau unei mici deletii se creeaza diversitatea cromozomilor, care este considerata o conditie necesara pentru evolutie.
Dacă apar recombinări între elementele mobile situate pe diferiți cromozomi, atunci se formează rearanjamente cromozomiale, care, în timpul diviziunilor celulare ulterioare, pot duce la un dezechilibru în genom. Iar un genom dezechilibrat, ca un buget dezechilibrat, este foarte prost divizat. Deci moartea genomilor nereușiți este unul dintre motivele pentru care elementele active transpozabile nu inundă cromozomii fără limită.
Apare o întrebare firească: cât de semnificativă este contribuția elementelor mobile la evoluție? În primul rând, majoritatea elementelor transpozabile sunt introduse, grosier vorbind, acolo unde trebuie, drept urmare pot deteriora sau modifica structura sau reglarea genei în care sunt introduse. Apoi selecția naturală mătură deoparte opțiunile nereușite, iar opțiunile de succes cu proprietăți adaptative sunt remediate.

Dacă consecințele introducerii unui element transposabil se dovedesc a fi neutre, atunci această variantă poate fi păstrată în populație, oferind o anumită diversitate în structura genei. Acest lucru poate fi util atunci când Condiții nefavorabile. Teoretic, în timpul mișcării în masă a elementelor mobile, pot apărea mutații în mai multe gene în același timp, ceea ce poate fi foarte util în cazul unei schimbări bruște a condițiilor de existență.
Deci, pentru a rezuma: există multe elemente mobile în genom și sunt diferite; pot interacționa atât între ele, cât și cu genele gazdă; poate fi dăunătoare și de neînlocuit. Instabilitatea genomului cauzată de mișcarea elementelor mobile se poate termina într-o tragedie pentru un individ, dar capacitatea de a se schimba rapid este o condiție necesară pentru supraviețuirea unei populații sau a unei specii. Aceasta creează diversitatea, care stă la baza selecției naturale și a transformărilor evolutive ulterioare.
Puteți face o analogie între genele săritoare și imigranți: unii imigranți sau descendenții lor devin cetățeni egali, altora li se acordă permise de ședere, iar alții - cei care nu respectă legile - sunt deportați sau închiși. Iar migrațiile în masă ale popoarelor pot schimba rapid statul în sine.
Literatură
Ratner V. A., Vasilyeva L. A. Inducerea transpozițiilor elementelor genetice mobile prin influențe stresante. Legatura ruseasca. 2000.
Gvozdev V. A. ADN eucariotic mobil // Soros Educational Journal. 1998. Nr. 8.





