Deshifrimi i gjenomit njerëzor. Gjenetika mjekësore Gjatë deshifrimit të gjenomit të Drosophila, u zbulua se
Për 50 vjetorin e zbulimit të strukturës së ADN-së
A.V. Zelenin
GJENOMI I BIMËVE
A. V. Zelenin
Zelenin Alexander Vladimirovich- d.b.n.,
shef i laboratorit të Institutit të Biologjisë Molekulare. V.A. Engelhardt RAS.
Arritjet mbresëlënëse të programit "Genomi i njeriut", si dhe suksesi i punës për deshifrimin e të ashtuquajturave gjenome ekstra të vogla (viruse), të vogla (baktere, maja) dhe të mesme (krimbët e rrumbullakët, Drosophila), bënë të mundur që të kaloni në një studim në shkallë të gjerë të gjenomave të bimëve të mëdha dhe tepër të mëdha. Nevoja urgjente për një studim të hollësishëm të gjenomave të bimëve më të rëndësishme ekonomikisht u theksua në një takim mbi gjenomikën e bimëve të mbajtur në vitin 1997 në Shtetet e Bashkuara [ , ]. Gjatë viteve që kanë kaluar që nga ajo kohë, janë arritur suksese të padyshimta në këtë fushë. Në vitin 2000, u shfaq një botim mbi sekuencën e plotë (përcaktimin e sekuencës nukleotide lineare të të gjithë ADN-së bërthamore) të gjenomit të mustardës së vogël - Arabidopsis, në 2001 - mbi sekuencën paraprake (draft) të gjenomit të orizit. Punimet për renditjen e gjenomave të bimëve të mëdha dhe super të mëdha (misër, thekër, grurë) u raportuan vazhdimisht, megjithatë, këto raporte nuk përmbanin informacion specifik dhe ishin më tepër në natyrën e deklaratave të qëllimit.
Supozohet se dekodimi i gjenomave të bimëve do të hapë perspektiva të gjera për shkencën dhe praktikën. Para së gjithash, identifikimi i gjeneve të reja dhe zinxhiri i rregullimit të tyre gjenetik do të rrisë ndjeshëm produktivitetin e bimëve nëpërmjet përdorimit të qasjeve bioteknologjike. Me zbulimin, izolimin, riprodhimin (klonimin) dhe renditjen e gjeneve përgjegjëse për funksione kaq të rëndësishme të organizmit bimor si riprodhimi dhe produktiviteti, proceset e ndryshueshmërisë, rezistenca ndaj faktorëve të pafavorshëm mjedisor, si dhe çiftimi homolog i kromozomeve, shfaqja e shoqërohen mundësi të reja për përmirësimin e procesit të mbarështimit. Së fundi, gjenet e izoluara dhe të klonuara mund të përdoren për të marrë bimë transgjenike me veti thelbësisht të reja dhe për të analizuar mekanizmat e rregullimit të aktivitetit të gjeneve.
Rëndësia e studimit të gjenomave të bimëve theksohet edhe nga fakti se deri tani numri i gjeneve të bimëve të lokalizuara, të klonuara dhe të sekuencave është i vogël dhe varion, sipas vlerësimeve të ndryshme, midis 800 dhe 1200. Kjo është 10-15 herë më pak se, për për shembull, te njerëzit.
Shtetet e Bashkuara mbeten lider i padyshimtë në studimin në shkallë të gjerë të gjenomave të bimëve, megjithëse studime intensive të gjenomit të orizit po kryhen në Japoni dhe në vitet e fundit dhe në Kinë. Në deshifrimin e gjenomit të Arabidopsis, përveç laboratorëve amerikanë, morën pjesë aktive edhe grupe kërkimore evropiane. Udhëheqja e dukshme e Shteteve të Bashkuara shkakton shqetësim serioz të shkencëtarëve evropianë, të cilin ata e shprehën qartë në një takim me titullin domethënës "Perspektivat për gjenomikën në epokën post-gjenomike", mbajtur në fund të vitit 2000 në Francë. Përparimi i shkencës amerikane në studimin e gjenomit të bimëve bujqësore dhe krijimin e formave të bimëve transgjenike, sipas shkencëtarëve evropianë, kërcënon se në një të ardhme jo shumë të largët (dy deri në pesë dekada), kur rritja e popullsisë do ta vendosë njerëzimin përballë një kriza ushqimore, ekonomia dhe shkenca evropiane do të varen nga teknologjia amerikane. Në këtë drejtim u njoftua krijimi i një programi shkencor franko-gjerman për studimin e gjenomave të bimëve (“Plantgene”) dhe në të u bënë investime të konsiderueshme.
Natyrisht, problemet e gjenomikës së bimëve duhet të tërheqin vëmendjen e ngushtë të shkencëtarëve dhe organizatorëve rusë të shkencës, si dhe të organeve drejtuese, pasi nuk ka të bëjë vetëm me prestigjin shkencor, por edhe me sigurinë kombëtare të vendit. Në një ose dy dekada, ushqimi do të bëhet burimi më i rëndësishëm strategjik.
VËSHTIRËSITË NË STUDIMIN E GJENOMËVE TË BIMËVE
Studimi i gjenomit të bimëve është një detyrë shumë më e vështirë sesa studimi i gjenomit të njerëzve dhe kafshëve të tjera. Kjo është për shkak të rrethanave të mëposhtme:
madhësive të mëdha të gjenomit, duke arritur për lloje të caktuara bimët me dhjetëra dhe madje qindra miliarda çifte bazash (bp): gjenomet e bimëve kryesore ekonomikisht të rëndësishme (përveç orizit, lirit dhe pambukut) janë ose të përmasave afër gjenomit njerëzor ose shumë herë më të mëdha (tabela);STUDIME KROMOZOMALE TË GJENOMEVELuhatje të mprehta në numrin e kromozomeve në bimë të ndryshme - nga dy në disa specie në disa qindra në të tjera, dhe nuk është e mundur të identifikohet një korrelacion i rreptë midis madhësisë së gjenomit dhe numrit të kromozomeve;
Një bollëk poliploidësh (që përmbajnë më shumë se dy gjenome për qelizë) me gjenome të ngjashme, por jo identike (alpolyploidy);
Pasurimi ekstrem i gjenomave të bimëve (deri në 99%) të ADN-së "të parëndësishme" (jo koduese, d.m.th., që nuk përmban gjene), e cila e ndërlikon shumë ngjitjen (rregullimin në rendin e duhur) të fragmenteve të sekuencave në një të përbashkët të gjerë. rajoni i ADN-së me madhësi (contig);
Harta morfologjike, gjenetike dhe fizike e kromozomeve jo të plota (në krahasim me gjenomin Drosophila, të njeriut dhe të miut);
Pamundësia praktike e izolimit të kromozomeve individuale në formë të pastër duke përdorur metoda që zakonisht përdoren për këtë qëllim për kromozomet e njeriut dhe të kafshëve (ndarja në një rrjedhë dhe përdorimi i hibrideve qelizore);
Vështirësia e hartës kromozomike (përcaktimi i vendndodhjes në kromozom) të gjeneve individuale duke përdorur hibridizimin në vend, si për shkak të përmbajtjes së lartë të ADN-së "të parëndësishme" në gjenomet e bimëve, ashtu edhe për veçoritë e organizimit strukturor të kromozomeve bimore;
Largësia evolucionare e bimëve nga kafshët, e cila ndërlikon seriozisht përdorimin e informacionit të marrë nga renditja e gjenomave të njerëzve dhe kafshëve të tjera për studimin e gjenomave të bimëve;
Procesi i gjatë i riprodhimit të shumicës së bimëve, i cili ngadalëson ndjeshëm analizën e tyre gjenetike.
Studimet kromozomale (citogjenetike) të gjenomave në përgjithësi dhe bimëve në veçanti kanë histori e gjatë. Termi "gjenom" u propozua për të treguar një grup haploid (të vetëm) kromozomesh me gjenet e përfshira në to në çerekun e parë të shekullit të 20-të, domethënë shumë kohë përpara vendosjes së rolit të ADN-së si bartës i informacionit gjenetik. .
Përshkrimi i gjenomit të një organizmi shumëqelizor të ri, të pastudiuar gjenetikisht më parë, zakonisht fillon me studimin dhe përshkrimin e grupit të plotë të kromozomeve të tij (kariotip). Kjo, natyrisht, vlen edhe për bimët, një numër i madh i të cilave as nuk kanë filluar të studiohen.
Tashmë në agimin e studimeve të kromozomeve, gjenomet e specieve bimore të lidhura u krahasuan bazuar në analizën e konjugimit mejotik (kombinimi i kromozomeve homologe) në hibridet ndërspecifike. Gjatë 100 viteve të fundit, mundësitë e analizës së kromozomeve janë zgjeruar në mënyrë dramatike. Tani, teknologjitë më të avancuara përdoren për të karakterizuar gjenomet e bimëve: variante të ndryshme të të ashtuquajturës ngjyrosje diferenciale, e cila bën të mundur identifikimin e kromozomeve individuale sipas veçorive morfologjike; hibridizimi në vend duke bërë të mundur lokalizimin e gjeneve specifike në kromozome; kërkime biokimike proteinat qelizore (elektroforeza dhe imunokimia) dhe, së fundi, një grup metodash të bazuara në analizën e ADN-së kromozomale deri në renditjen e saj.

Oriz. një. Kariotipet e drithërave a - thekra (14 kromozome), b - gruri i fortë (28 kromozome), c - gruri i butë (42 kromozome), d - elbi (14 kromozome)Për shumë vite, janë studiuar kariotipet e drithërave, kryesisht gruri dhe thekra. Është interesante që tipe te ndryshme këto bimë, numri i kromozomeve është i ndryshëm, por gjithmonë shumëfish i shtatë. Llojet individuale të drithërave mund të njihen me besueshmëri nga kariotipi i tyre. Për shembull, gjenomi i thekrës përbëhet nga shtatë çifte kromozomesh të mëdhenj me blloqe heterokromatike me ngjyra intensive në skajet e tyre, shpesh të quajtura segmente ose breza (Fig. 1a). Gjenomet e grurit kanë tashmë 14 dhe 21 çifte kromozomesh (Fig. 1, b, c), dhe shpërndarja e blloqeve heterokromatike në to nuk është e njëjtë si në kromozomet e thekrës. Gjenomet individuale të grurit, të emërtuara A, B dhe D, gjithashtu ndryshojnë nga njëra-tjetra. Një rritje në numrin e kromozomeve nga 14 në 21 çon në një ndryshim të mprehtë në vetitë e grurit, gjë që pasqyrohet në emrat e tyre: durum, ose makarona, grurë dhe të buta, ose bukë, grurë . Gjeni D, i cili përmban gjene për proteinat e glutenit, i cili i jep brumit të ashtuquajturin mbirje, është përgjegjës për përvetësimin e vetive të larta të pjekjes nga gruri i butë. Pikërisht këtij gjenomi i kushtohet vëmendje e veçantë në përmirësimin e përzgjedhjes së grurit të bukës. Një tjetër drithëra me 14 kromozome, elbi (Fig. 1, d), zakonisht nuk përdoret për të bërë bukë, por është lënda e parë kryesore për prodhimin e produkteve të tilla të zakonshme si birra dhe uiski.
Po studiohen intensivisht kromozomet e disa bimëve të egra që përdoren për të përmirësuar cilësinë e specieve më të rëndësishme bujqësore, siç janë të afërmit e egër të grurit - Aegilops. Format e reja bimore krijohen nga kryqëzimi (Fig. 2) dhe seleksionimi. Vitet e fundit, një përmirësim i ndjeshëm i metodave të kërkimit ka bërë të mundur fillimin e studimit të gjenomave të bimëve, tiparet e kariotipeve të të cilave (kryesisht madhësia e vogël e kromozomeve) i bënë ato më parë të paarritshme për analizën e kromozomeve. Pra, vetëm kohët e fundit të gjitha kromozomet e pambukut, kamomilit dhe lirit u identifikuan për herë të parë.

Oriz. 2. Kariotipet e grurit dhe një hibrid i grurit me Aegilops
a - grurë i butë heksaploid ( Triticum astivum), i përbërë nga gjenomet A, B dhe O; b - grurë tetraploid ( Triticum timopheevi), i përbërë nga gjenomet A dhe G. përmban gjene për rezistencë ndaj shumicës së sëmundjeve të grurit; c - hibride Triticum astivum X Triticum timopheevi rezistent ndaj mykut pluhur dhe ndryshkut, zëvendësimi i një pjese të kromozomeve është qartë i dukshëmSTRUKTURA PRIMARE E ADN-së
Me zhvillimin e gjenetikës molekulare, vetë koncepti i gjenomit është zgjeruar. Tani ky term interpretohet si në kromozomin klasik ashtu edhe në kuptimin molekular modern: i gjithë materiali gjenetik i një virusi, qelize dhe organizmi individual. Natyrisht, pas studimit të strukturës së plotë parësore të gjenomave (siç quhet shpesh sekuenca e plotë lineare e bazave të acidit nukleik) të një numri mikroorganizmash dhe njerëzve, u ngrit çështja e renditjes së gjenomit bimor.
Nga shumë organizma bimorë, dy u zgjodhën për studim - Arabidopsis, që përfaqëson klasën e dikoteve (madhësia e gjenomit 125 milion bp) dhe orizi nga klasa e monocots (420-470 milion bp). Këto gjenome janë të vogla në krahasim me gjenomet e tjera bimore dhe përmbajnë relativisht pak segmente të përsëritura të ADN-së. Veçori të tilla dhanë shpresë se gjenomet e përzgjedhura do të ishin të disponueshme për përcaktimin relativisht të shpejtë të strukturës së tyre parësore.

Oriz. 3. Arabidopsis - mustardë e vogël - një bimë e vogël nga familja e kryqëzatave ( Brassicaceae). Në një hapësirë të barabartë në sipërfaqe me një faqe të revistës sonë, ju mund të rritni deri në një mijë organizma individualë Arabidopsis.Arsyeja e zgjedhjes së Arabidopsis nuk ishte vetëm madhësia e vogël e gjenomit të saj, por edhe madhësia e vogël e organizmit, gjë që e bën të lehtë rritjen e saj në laborator (Fig. 3). Ne morëm parasysh ciklin e tij të shkurtër riprodhues, falë të cilit është e mundur të kryhen shpejt eksperimente për kryqëzimin dhe përzgjedhjen, gjenetikën e studiuar në detaje, lehtësinë e manipulimit me ndryshimin e kushteve të rritjes (ndryshimi i përbërjes së kripës së tokës, shtimi i lëndëve ushqyese të ndryshme, etj. .) dhe testimi i efektit në bimë të faktorëve dhe patogjenëve të ndryshëm mutagjenë (viruse, baktere, kërpudha). Arabidopsis nuk ka asnjë vlerë ekonomike, prandaj gjenomi i tij, së bashku me gjenomin e miut, u quajt referencë, ose, më pak saktë, model.*
* Shfaqja e termit "gjenom model" në literaturën ruse është rezultat i një përkthimi të pasaktë të frazës angleze model genome. Fjala "model" nënkupton jo vetëm mbiemrin "model", por edhe emrin "kampion", "standard", "model". Do të ishte më e saktë të flitej për një gjenom mostër, ose një gjenom referencë.Puna intensive për renditjen e gjenomit të Arabidopsis filloi në vitin 1996 nga një konsorcium ndërkombëtar që përfshinte institucione shkencore dhe grupe kërkimore nga SHBA, Japonia, Belgjika, Italia, Britania e Madhe dhe Gjermania. Në dhjetor 2000, informacione të gjera u vunë në dispozicion duke përmbledhur përcaktimin e strukturës parësore të gjenomit Arabidopsis. Për renditjen u përdor teknologjia klasike ose hierarkike: së pari, u studiuan seksione të vogla individuale të gjenomit, nga të cilat përbëheshin seksione më të mëdha (contigs) dhe, në fazën përfundimtare, struktura e kromozomeve individuale. ADN-ja bërthamore e gjenomit Arabidopsis shpërndahet në pesë kromozome. Në vitin 1999, u publikuan rezultatet e sekuencës së dy kromozomeve dhe shfaqja në shtyp e informacionit në lidhje me strukturën parësore të tre të tjerëve përfundoi sekuencën e të gjithë gjenomit.
Nga 125 milionë çifte bazash, është përcaktuar struktura primare prej 119 milionë, që është 92% e të gjithë gjenomit. Vetëm 8% e gjenomit Arabidopsis që përmban blloqe të mëdha të segmenteve të përsëritura të ADN-së doli të ishte i paarritshëm për studim. Për sa i përket tërësisë dhe plotësimit të sekuencës së gjenomit eukariotik, Arabidopsis mbetet në tre kampionët kryesorë së bashku me një organizëm majaje njëqelizore. Saccharomyces cerevisiae dhe organizmit shumëqelizor Eleganca e Caenorhabditis(shih tabelën).
Rreth 15,000 gjene individuale të kodimit të proteinave janë gjetur në gjenomën Arabidopsis. Përafërsisht 12,000 prej tyre përmbahen si dy kopje për gjenom haploid (të vetëm), kështu që numri total gjenet janë 27 mijë. Numri i gjeneve në Arabidopsis nuk ndryshon shumë nga numri i gjeneve në organizmat si njerëzit dhe minjtë, por madhësia e gjenomit të tij është 25-30 herë më e vogël. Kjo rrethanë lidhet me karakteristika të rëndësishme në strukturën e gjeneve individuale të Arabidopsis dhe strukturën e përgjithshme të gjenomit të tij.
Gjenet e Arabidopsis janë kompakte, që përmbajnë vetëm disa ekzone (rajone koduese të proteinave) të ndara nga segmente të shkurtra (rreth 250 bp) jo-koduese të ADN-së (introne). Intervalet ndërmjet gjeneve individuale janë mesatarisht 4600 çifte bazash. Për krahasim, theksojmë se gjenet njerëzore përmbajnë shumë dhjetëra dhe madje qindra ekzone dhe introne, dhe rajonet ndërgjenike kanë madhësi prej 10 mijë çifte bazash ose më shumë. Supozohet se prania e një gjenomi të vogël kompakt kontribuoi në stabilitetin evolucionar të Arabidopsis, pasi ADN-ja e tij u bë një objektiv për agjentë të ndryshëm dëmtues në një masë më të vogël, në veçanti, për futjen e fragmenteve të ADN-së të ngjashme me virusin (transpozonët). në gjenom.
Ndër veçoritë e tjera molekulare të gjenomit Arabidopsis, duhet theksuar se ekzonet pasurohen me guaninë dhe citozinë (44% në ekzone dhe 32% në introne) krahasuar me gjenet e kafshëve, si dhe prania e gjeneve të përsëritura dyfish (të dyfishuara). Supozohet se një dyfishim i tillë ka ndodhur si rezultat i katër ngjarjeve të njëkohshme, që konsistojnë në dyfishimin (përsëritjen) e një pjese të gjeneve Arabidopsis, ose shkrirjen e gjenomave të lidhura me to. Këto ngjarje, të ndodhura 100-200 milion vjet më parë, janë një manifestim i prirjes së përgjithshme drejt poliploidizimit (rritje e shumëfishtë e numrit të gjenomave në një organizëm), që është karakteristikë e gjenomave të bimëve. Megjithatë, disa fakte tregojnë se gjenet e dyfishta në Arabidopsis nuk janë identike dhe funksionojnë ndryshe, gjë që mund të shoqërohet me mutacione në rajonet e tyre rregullatore.
Orizi është bërë një tjetër objekt i sekuencës së plotë të ADN-së. Gjenomi i kësaj bime është gjithashtu i vogël (12 kromozome, që japin një total prej 420-470 milion bp), vetëm 3,5 herë më i madh se ai i Arabidopsis. Sidoqoftë, ndryshe nga Arabidopsis, orizi ka një rëndësi të madhe ekonomike, duke qenë baza e të ushqyerit për më shumë se gjysmën e njerëzimit, prandaj, jo vetëm miliarda konsumatorë, por edhe një ushtri shumëmilionëshe njerëzish të përfshirë në mënyrë aktive në procesin shumë të mundimshëm të tij. kultivimi janë jashtëzakonisht të interesuar në përmirësimin e vetive të tij.
Disa studiues filluan të studiojnë gjenomën e orizit që në vitet 1980, por këto studime arritën një shkallë serioze vetëm në vitet 1990. Në vitin 1991, në Japoni u krijua një program për të deshifruar strukturën e gjenomit të orizit, duke bashkuar përpjekjet e shumë grupeve kërkimore. Në vitin 1997, mbi bazën e këtij programi u organizua Projekti Ndërkombëtar i Gjenomit të Orizit. Pjesëmarrësit e tij vendosën të përqendrojnë përpjekjet e tyre në renditjen e një prej nënspecieve të orizit ( Oriza sativajaponica), në studimin e së cilës ishte arritur tashmë përparim i rëndësishëm deri në atë kohë. Një stimul serioz dhe, në mënyrë figurative, një yll udhëzues për një punë të tillë ishte programi "Gjenomi i njeriut".
Në kuadër të këtij programi u testua strategjia e ndarjes hierarkike “kromozomale” të gjenomit, të cilën pjesëmarrësit e konsorciumit ndërkombëtar e përdorën për deshifrimin e gjenomit të orizit. Sidoqoftë, nëse në studimin e gjenomit njerëzor, fraksionet e kromozomeve individuale u izoluan duke përdorur metoda të ndryshme, atëherë materiali specifik për kromozomet individuale të orizit dhe rajonet e tyre individuale u mor me mikrodiseksion lazer (prerja e objekteve mikroskopike). Në një rrëshqitje mikroskopi, ku ndodhen kromozomet e orizit, nën ndikimin e një rreze lazer, gjithçka digjet, përveç kromozomit ose pjesëve të tij të planifikuara për analizë. Materiali i mbetur përdoret për klonim dhe sekuencë.
Janë publikuar raporte të shumta mbi rezultatet e renditjes së fragmenteve individuale të gjenomës së orizit, të kryera me saktësi dhe detaje të larta, karakteristikë e teknologjisë hierarkike. Besohej se përcaktimi i strukturës së plotë parësore të gjenomit të orizit do të përfundonte në fund të vitit 2003-mesi i vitit 2004 dhe rezultatet, së bashku me të dhënat mbi strukturën parësore të gjenomit Arabidopsis, do të përdoreshin gjerësisht në krahasimin. gjenomika e bimëve të tjera.
Megjithatë, në fillim të vitit 2002, dy grupe kërkimore - njëri nga Kina, tjetri nga Zvicra dhe Shtetet e Bashkuara - publikuan rezultatet e një drafti të plotë (të përafërt) të sekuencës së gjenomit të orizit, të kryer duke përdorur teknologjinë e klonimit total. Në ndryshim nga studimi i staduar (hierarkik), qasja totale bazohet në klonimin e njëkohshëm të të gjithë ADN-së gjenomike në një nga vektorët viralë ose bakterialë dhe marrjen e një numri të konsiderueshëm (të madh për gjenomet e mesme dhe të mëdha) të kloneve individuale që përmbajnë të ndryshëm Segmentet e ADN-së. Bazuar në analizën e këtyre seksioneve të sekuencave dhe mbivendosjes së seksioneve identike terminale të ADN-së, formohet një kontig - një zinxhir sekuencash të ADN-së të bashkuara së bashku. Kontigu i përgjithshëm (gjithsej) është struktura primare e të gjithë gjenomit, ose të paktën e një kromozomi individual.
Në një paraqitje të tillë skematike, strategjia e klonimit total duket e thjeshtë. Në fakt, ai ndeshet me vështirësi serioze që lidhen me nevojën për të marrë një numër të madh klonesh (përgjithësisht pranohet që gjenomi ose rajoni i tij në studim duhet të mbivendosen nga klone të paktën 10 herë), sasinë e madhe të sekuencave dhe jashtëzakonisht punë komplekse e kloneve docking që kërkojnë pjesëmarrjen e specialistëve të bioinformatikës. Një pengesë serioze për klonimin total është një shumëllojshmëri segmentesh të përsëritura të ADN-së, numri i të cilave, siç u përmend tashmë, rritet ndjeshëm me rritjen e madhësisë së gjenomit. Prandaj, strategjia e sekuencës totale përdoret kryesisht në studimin e gjenomave të viruseve dhe mikroorganizmave, megjithëse është përdorur me sukses për të studiuar gjenomin e një organizmi shumëqelizor, Drosophila.
Rezultatet e sekuencës totale të këtij gjenomi u "mbivendosën" në një grup të madh informacioni rreth strukturës së tij kromozomale, gjenesh dhe molekulare të marra gjatë një periudhe pothuajse 100-vjeçare të studimit të Drosophila. E megjithatë, për sa i përket shkallës së renditjes, gjenomi Drosophila (66% e madhësisë totale të gjenomit) është dukshëm inferior ndaj gjenomit të Arabidopsis (92%), megjithë madhësitë e tyre mjaft të afërta - përkatësisht 180 milion dhe 125 milion çifte bazë. . Prandaj, kohët e fundit është propozuar të emërtohet teknologjia e përzier, e cila u përdor për sekuencimin e gjenomit Drosophila.
Për të renditur gjenomin e orizit, grupet kërkimore të përmendura më sipër morën dy nga nënllojet e tij, më të kultivuarat në vendet aziatike, - Oriza saliva L. ssp indicaj dhe Oriza saliva L. sspjaponica. Rezultatet e studimeve të tyre përkojnë në shumë aspekte, por ndryshojnë në shumë aspekte. Kështu, përfaqësuesit e të dy grupeve deklaruan se kishin arritur afërsisht 92-93% të mbivendosjes së gjenomit me kontigs. Është treguar se rreth 42% e gjenomës së orizit përfaqësohet nga përsëritje të shkurtra të ADN-së që përbëhen nga 20 çifte bazash, dhe shumica e elementëve të lëvizshëm të ADN-së (transpozonëve) ndodhen në rajone ndërgjenike. Megjithatë, të dhënat për madhësinë e gjenomit të orizit ndryshojnë ndjeshëm.
Për nëngrupin japonez, madhësia e gjenomit është përcaktuar të jetë 466 milionë çifte bazë, dhe për nëngrupin indian, 420 milionë. Arsyeja e kësaj mospërputhjeje nuk është e qartë. Mund të jetë pasojë e qasjeve të ndryshme metodologjike në përcaktimin e madhësisë së pjesës jokoduese të gjenomit, domethënë nuk pasqyron gjendjen e vërtetë të punëve. Por është e mundur që një ndryshim prej 15% në madhësinë e gjenomave të studiuara ekziston.
Mospërputhja e dytë e madhe u zbulua në numrin e gjeneve të gjetura: për nëngrupin japonez, nga 46,022 në 55,615 gjene për gjenom, dhe për nënllojet indiane, nga 32,000 në 50,000. Arsyeja për këtë mospërputhje nuk është e qartë.
Paplotësia dhe mospërputhja e informacionit të marrë vërehet në komentet e artikujve të botuar. Shpresa shprehet gjithashtu këtu se boshllëqet në njohuritë e gjenomit të orizit do të eliminohen duke krahasuar të dhënat e "sekuencës së përafërt" me rezultatet e renditjes së detajuar hierarkike të kryer nga pjesëmarrësit e Projektit Ndërkombëtar të Gjenomit të Orizit.
GJENOMIA KRAHASUESE DHE FUNKSIONALE E BIMËVE
Të dhënat e gjera të marra, gjysma e të cilave (rezultatet e grupit kinez) janë të disponueshme publikisht, hapin padyshim perspektiva të gjera si për studimin e gjenomit të orizit ashtu edhe për gjenomikën e bimëve në përgjithësi. Krahasimi i vetive të gjenomit të Arabidopsis dhe orizit tregoi se shumica e gjeneve (deri në 80%) të identifikuara në gjenomin Arabidopsis gjenden gjithashtu në gjenomën e orizit, megjithatë, për afërsisht gjysmën e gjeneve të gjetura në oriz, analoge (ortologë ) nuk janë gjetur ende në gjenomin Arabidopsis. . Në të njëjtën kohë, 98% e gjeneve, struktura primare e të cilave është krijuar për drithëra të tjera, u gjetën në gjenomën e orizit.
Mospërputhja e rëndësishme (pothuajse e dyfishtë) midis numrit të gjeneve në oriz dhe Arabidopsis është e çuditshme. Në të njëjtën kohë, të dhënat e draftit të dekodimit të gjenomës së orizit, të marra duke përdorur sekuencën totale, praktikisht nuk krahasohen me rezultatet e gjera të studimit të gjenomit të orizit me metodën e klonimit dhe sekuencës hierarkike, domethënë atë që ka është bërë në lidhje me gjenomin Drosophila nuk është kryer. Prandaj, mbetet e paqartë nëse ndryshimi në numrin e gjeneve në Arabidopsis dhe oriz pasqyron gjendjen e vërtetë të punëve apo nëse shpjegohet me ndryshimin në qasjet metodologjike.
Në ndryshim nga gjenomi i Arabidopsis, të dhënat për gjenet binjake në gjenomën e orizit nuk janë dhënë. Është e mundur që sasia e tyre relative të jetë më e lartë në oriz sesa në Arabidopsis. Kjo mundësi mbështetet në mënyrë indirekte nga të dhënat për praninë e formave poliploide të orizit. Më shumë qartësi për këtë çështje mund të pritet pasi të përfundojë Projekti Ndërkombëtar i Gjenomit të Orizit dhe të merret një pamje e detajuar e strukturës primare të ADN-së të këtij gjenomi. Baza serioze për një shpresë të tillë jep fakti se pas botimit të punimeve mbi sekuencën e përafërt të gjenomit të orizit, numri i botimeve mbi strukturën e këtij gjenomi është rritur ndjeshëm, në veçanti, informacioni është shfaqur për renditjen e detajuar. të kromozomeve 1 dhe 4 të tij.
Njohja, të paktën përafërsisht, e numrit të gjeneve në bimë është me rëndësi themelore për gjenomikën krahasuese të bimëve. Fillimisht, besohej se duke qenë se të gjitha bimët e lulëzuara janë shumë afër njëra-tjetrës për sa i përket karakteristikave të tyre fenotipike, gjenomet e tyre gjithashtu duhet të jenë të ngjashme. Dhe nëse studiojmë gjenomin e Arabidopsis, do të marrim informacion për shumicën e gjenomave të bimëve të tjera. Një konfirmim indirekt i këtij supozimi janë rezultatet e sekuencës së gjenomit të miut, i cili është çuditërisht afër gjenomit njerëzor (rreth 30 mijë gjene, nga të cilat vetëm 1 mijë rezultuan të jenë të ndryshëm).
Mund të supozohet se arsyeja e ndryshimeve midis gjenomave të Arabidopsis dhe orizit qëndron në përkatësinë e tyre ndaj klasave të ndryshme të bimëve - dikot dhe monokotë. Për të sqaruar këtë çështje, është shumë e dëshirueshme të njihni të paktën një strukturë primare të përafërt të ndonjë bime tjetër monokotiledone. Kandidati më real mund të jetë misri, gjenomi i të cilit është afërsisht i barabartë me gjenomin e njeriut, por gjithsesi shumë më i vogël se gjenomi i drithërave të tjera. Vlera ushqyese e misrit është e njohur mirë.
Materiali i madh i marrë si rezultat i sekuencës së gjenomit të Arabidopsis dhe orizit po bëhet gradualisht baza për një studim në shkallë të gjerë të gjenomave të bimëve duke përdorur gjenomikën krahasuese. Studime të tilla kanë një rëndësi të përgjithshme biologjike, pasi ato na lejojnë të vendosim parimet kryesore të organizimit të gjenomit të bimëve në tërësi dhe kromozomeve të tyre individuale, për të identifikuar tipare të përbashkëta strukturën e gjeneve dhe rajonet e tyre rregullatore, për të marrë në konsideratë raportin e pjesës (gjenit) funksionalisht aktive të kromozomit dhe rajoneve të ndryshme ndërgjenike të ADN-së që nuk kodojnë për proteinat. Gjenetika krahasuese po bëhet gjithashtu gjithnjë e më e rëndësishme për zhvillimin e gjenomikës funksionale njerëzore. Pikërisht për studime krahasuese u krye sekuenca e gjenomit të peshkut puffer dhe të miut.
Po aq i rëndësishëm është studimi i gjeneve individuale përgjegjëse për sintezën e proteinave individuale që përcaktojnë funksione specifike të trupit. Pikërisht në zbulimin, izolimin, renditjen dhe përcaktimin e funksionit të gjeneve individuale qëndron rëndësia praktike, kryesisht mjekësore, e programit të Gjenomit Njerëzor. Kjo rrethanë u vu në dukje disa vite më parë nga J. Watson, i cili theksoi se programi "Gjenomi i njeriut" do të përfundonte vetëm kur të përcaktoheshin funksionet e të gjitha gjeneve njerëzore.

Oriz. katër. Klasifikimi sipas funksionit të gjeneve të Arabidopsis
1 - gjenet për rritjen, ndarjen dhe sintezën e ADN-së; 2 - gjenet e sintezës së ARN-së (transkriptimi); 3 - gjenet për sintezën dhe modifikimin e proteinave; 4 - gjenet për zhvillimin, plakjen dhe vdekjen e qelizave; 5 - gjenet e metabolizmit qelizor dhe metabolizmit të energjisë; 6 - gjenet e ndërveprimit ndërqelizor dhe transmetimit të sinjalit; 7 - gjenet për sigurimin e proceseve të tjera qelizore; 8 - gjene me funksion të panjohurSa i përket funksionit të gjeneve të bimëve, ne dimë më pak se një të dhjetën e asaj që dimë për gjenet njerëzore. Edhe në Arabidopsis, gjenomi i të cilit është shumë më i studiuar se gjenomi i njeriut, funksioni i pothuajse gjysmës së gjeneve të tij mbetet i panjohur (Fig. 4). Ndërkohë, përveç gjeneve të përbashkëta me kafshët, bimët kanë një numër të konsiderueshëm gjenesh që janë specifike vetëm (ose të paktën kryesisht) për to. Bëhet fjalë për për gjenet që marrin pjesë në transportin e ujit dhe sintezën e murit qelizor, i cili mungon te kafshët, për gjenet që sigurojnë formimin dhe funksionimin e kloroplasteve, fotosintezën, fiksimin e azotit dhe sintezën e produkteve të shumta aromatike. Kjo listë mund të vazhdojë, por tashmë është e qartë se çfarë detyre të vështirë përballet gjenomika funksionale e bimëve.
Sekuenca e plotë e gjenomit siguron informacion afërsisht të vërtetë për numrin total të gjeneve në një organizëm të caktuar, bën të mundur vendosjen e informacionit pak a shumë të detajuar dhe të besueshëm për strukturën e tyre në bankat e të dhënave dhe lehtëson punën e izolimit dhe studimit të gjeneve individuale. Megjithatë, sekuenca e gjenomit nuk do të thotë në asnjë mënyrë vendosjen e funksionit të të gjitha gjeneve.
Një nga qasjet më premtuese të gjenomikës funksionale bazohet në identifikimin e gjeneve të punës që përdoren për transkriptimin (leximin) e mARN-së. Kjo qasje, duke përfshirë Teknologji moderne microarrays, ju lejon të identifikoni në të njëjtën kohë deri në dhjetëra mijëra gjene funksionale. Kohët e fundit, duke përdorur këtë qasje, ka filluar studimi i gjenomave të bimëve. Për Arabidopsis, ishte e mundur të merreshin rreth 26 mijë transkripte individuale, gjë që lehtëson shumë mundësinë e përcaktimit të funksionit të pothuajse të gjithë gjeneve të tij. Në patate, ishte e mundur të identifikoheshin rreth 20,000 gjene funksionale që janë të rëndësishme për të kuptuar si proceset e rritjes dhe formimit të tuberit, ashtu edhe proceset e sëmundjes së patates. Supozohet se kjo njohuri do të rrisë rezistencën e një prej produkteve ushqimore më të rëndësishme ndaj patogjenëve.
Zhvillimi logjik i gjenomikës funksionale ishte proteomika. Kjo fushë e re e shkencës studion proteomën, e cila zakonisht kuptohet si grupi i plotë i proteinave në një qelizë në një moment të caktuar. Një grup i tillë proteinash, duke pasqyruar gjendjen funksionale të gjenomit, ndryshon gjatë gjithë kohës, ndërsa gjenomi mbetet i pandryshuar.
Studimi i proteinave është përdorur prej kohësh për të gjykuar aktivitetin e gjenomave të bimëve. Siç dihet, enzimat e pranishme në të gjitha bimët ndryshojnë në specie dhe varietete individuale në sekuencën e aminoacideve. Enzima të tilla, me të njëjtin funksion, por një sekuencë të ndryshme të aminoacideve individuale, quhen izoenzima. Ato kanë veti të ndryshme fiziko-kimike dhe imunologjike (pesha molekulare, ngarkesa), të cilat mund të zbulohen duke përdorur kromatografi ose elektroforezë. Për shumë vite, këto metoda janë përdorur me sukses për të studiuar të ashtuquajturin polimorfizëm gjenetik, domethënë ndryshimet midis organizmave, varieteteve, popullatave, specieve, në veçanti grurit dhe formave të ngjashme të drithërave. Megjithatë, kohët e fundit, për shkak të zhvillimit të shpejtë të metodave të analizës së ADN-së, duke përfshirë sekuencën, studimi i polimorfizmit të proteinave është zëvendësuar nga studimi i polimorfizmit të ADN-së. Megjithatë, një metodë e rëndësishme dhe e besueshme për analizën gjenetike, përzgjedhjen dhe prodhimin e farës së bimëve bujqësore, mbetet studimi i drejtpërdrejtë i spektrave të proteinave ruajtëse (prolamina, gliadina, etj.), të cilat përcaktojnë vetitë kryesore ushqyese të drithërave.
Njohja e gjeneve, mekanizmave të shprehjes dhe rregullimit të tyre është jashtëzakonisht e rëndësishme për zhvillimin e bioteknologjisë dhe prodhimin e bimëve transgjenike. Dihet se sukseset mbresëlënëse në këtë fushë shkaktojnë një reagim të paqartë nga komuniteti mjedisor dhe mjekësor. Sidoqoftë, ekziston një fushë e bioteknologjisë së bimëve ku këto frikë, nëse jo plotësisht të pabaza, atëherë, në çdo rast, duket se kanë pak rëndësi. Po flasim për krijimin e impianteve industriale transgjenike që nuk përdoren si produkte ushqimore. India së fundmi korri prodhimin e parë të pambukut transgjenik që është rezistent ndaj një sërë sëmundjesh. Ka informacione për futjen e gjeneve speciale që kodojnë proteinat e pigmentit në gjenomën e pambukut dhe prodhimin e fibrave të pambukut që nuk kërkojnë ngjyrosje artificiale. Një tjetër kulturë industriale që mund të jetë objekt i inxhinierisë gjenetike efektive është liri. Përdorimi i tij si një alternativë ndaj pambukut për lëndët e para të tekstilit është diskutuar kohët e fundit. Ky problem është jashtëzakonisht i rëndësishëm për vendin tonë, i cili ka humbur burimet e veta të pambukut të papërpunuar.
PERSPEKTIVAT PËR STUDIMIN E GJENOMËVE TË BIMËVE
Natyrisht, studimet strukturore të gjenomave të bimëve do të bazohen në qasjet dhe metodat e gjenomikës krahasuese, duke përdorur si material kryesor rezultatet e deshifrimit të gjenomave të Arabidopsis dhe orizit. Një rol të rëndësishëm në zhvillimin e gjenomikës krahasuese të bimëve do të luajë padyshim informacioni që herët a vonë do të sigurohet nga sekuenca totale (e përafërt) e gjenomave të bimëve të tjera. Në këtë rast, gjenomika krahasuese e bimëve do të bazohet në vendosjen e marrëdhënieve gjenetike midis lokacioneve individuale dhe kromozomeve që i përkasin gjenomave të ndryshme. Ne do të fokusohemi jo aq në gjenomikën e përgjithshme të bimëve, sa në gjenomikën selektive të lokacioneve individuale kromozomale. Për shembull, kohët e fundit është treguar se gjeni përgjegjës për vernalizimin ndodhet në vendndodhjen VRn-AI të kromozomit hexaploid të grurit 5A dhe vendndodhjen Hd-6 të kromozomit 3 të orizit.
Zhvillimi i këtyre studimeve do të jetë një shtysë e fuqishme për identifikimin, izolimin dhe renditjen e shumë gjeneve të bimëve funksionalisht të rëndësishme, në veçanti, gjeneve përgjegjëse për rezistencën ndaj sëmundjeve, rezistencën ndaj thatësirës dhe përshtatshmërinë ndaj kushteve të ndryshme të rritjes. Gjithnjë e më shumë do të përdoret gjenomika funksionale, bazuar në zbulimin masiv (screening) të gjeneve që funksionojnë në bimë.
Mund të parashikojmë përmirësime të mëtejshme të teknologjive kromozomale, kryesisht metodën e mikrodiseksionit. Përdorimi i tij zgjeron në mënyrë dramatike mundësitë e kërkimit gjenomik pa kërkuar kosto të mëdha, siç është, për shembull, sekuenca totale e gjenomit. Metoda e lokalizimit në kromozomet e bimëve të gjeneve individuale me ndihmën e hibridizimit do të përhapet më tej. në vend. Për momentin, aplikimi i tij është i kufizuar nga numri i madh i sekuencave të përsëritura në gjenomën e bimëve, dhe ndoshta nga veçoritë e organizimit strukturor të kromozomeve bimore.
Teknologjitë kromozomale do të kenë një rëndësi të madhe për gjenominë evolucionare të bimëve në të ardhmen e parashikueshme. Këto teknologji relativisht të lira bëjnë të mundur vlerësimin e shpejtë të ndryshueshmërisë brenda dhe ndërspecifike, studimin e gjenomave komplekse alopoliploide të grurit tetraploid dhe hexaploid, triticale; të analizojë proceset evolucionare në nivel kromozomik; të hetojë formimin e gjenomave sintetike dhe futjen (introgresionin) e materialit gjenetik të huaj; identifikojnë marrëdhëniet gjenetike midis kromozomeve individuale lloje te ndryshme.
Studimi i kariotipit të bimës duke përdorur metoda klasike citogjenetike, të pasuruara me analiza biologjike molekulare dhe teknologji kompjuterike, do të përdoret për të karakterizuar gjenomin. Kjo është veçanërisht e rëndësishme për studimin e stabilitetit dhe ndryshueshmërisë së kariotipit në nivel jo vetëm të organizmave individualë, por edhe të popullatave, varieteteve dhe specieve. Së fundi, është e vështirë të imagjinohet se si mund të vlerësohet numri dhe spektri i rirregullimeve kromozomale (aberracionet, urat) pa përdorimin e metodave të ngjyrosjes diferenciale. Studime të tilla janë jashtëzakonisht premtuese për monitorimin e mjedisit sipas gjendjes së gjenomit të bimëve.
AT Rusia moderne sekuenca e drejtpërdrejtë e gjenomave të bimëve nuk ka gjasa. Një punë e tillë, që kërkon investime të mëdha, është përtej fuqisë së ekonomisë sonë aktuale. Ndërkohë, të dhënat për strukturën e gjenomave të Arabidopsis dhe orizit, të marra nga shkenca botërore dhe të disponueshme në bankat ndërkombëtare të të dhënave, janë të mjaftueshme për zhvillimin e gjenomikës së bimëve vendase. Mund të parashikohet zgjerimi i studimeve të gjenomave të bimëve bazuar në qasjet e gjenomikës krahasuese për zgjidhjen e problemeve specifike të mbarështimit dhe prodhimit të bimëve, si dhe për të studiuar origjinën e llojeve të ndryshme bimore me rëndësi të madhe ekonomike.
Mund të supozohet se qasjet gjenomike si tipizimi gjenetik (RELF, RAPD, analizat AFLP, etj.), të cilat janë mjaft të përballueshme për buxhetin tonë, do të përdoren gjerësisht në praktikën e mbarështimit vendas dhe në prodhimin e bimëve. Paralelisht me metodat e drejtpërdrejta për përcaktimin e polimorfizmit të ADN-së, qasjet e bazuara në studimin e polimorfizmit të proteinave, kryesisht proteinat ruajtëse të drithërave, do të përdoren për të zgjidhur problemet e gjenetikës dhe mbarështimit të bimëve. Teknologjitë kromozomale do të përdoren gjerësisht. Ato janë relativisht të lira, zhvillimi i tyre kërkon investime mjaft të moderuara. Në fushën e studimeve të kromozomeve, shkenca vendase nuk është inferiore ndaj botës.
Duhet theksuar se shkenca jonë ka dhënë një kontribut të rëndësishëm në formimin dhe zhvillimin e gjenomikës së bimëve [ , ].
Rolin themelor e luajti N.I. Vavilov (1887-1943).
Në biologjinë molekulare dhe gjenomikën e bimëve, kontributi pionier i A.N. Belozersky (1905-1972).
Në fushën e studimeve kromozomale, është e nevojshme të theksohet puna e gjenetistit të shquar S.G. Navashin (1857-1930), i cili së pari zbuloi kromozomet satelitore në bimë dhe vërtetoi se është e mundur të bëhet dallimi midis kromozomeve individuale sipas veçorive të morfologjisë së tyre.
Një tjetër klasik Shkenca ruse G.A. Levitsky (1878-1942) përshkroi në detaje kromozomet e thekrës, grurit, elbit, bizeles dhe panxharit të sheqerit, futi termin "kariotip" në shkencë dhe zhvilloi doktrinën e tij.
Specialistët modernë, duke u mbështetur në arritjet e shkencës botërore, mund të japin një kontribut të rëndësishëm në zhvillimin e mëtejshëm të gjenetikës dhe gjenomisë së bimëve.
Autori shpreh falënderimet e tij të përzemërta për Akademik Yu.P. Altukhov për diskutimin kritik të artikullit dhe këshilla të vlefshme.Puna e ekipit të kryesuar nga autori i artikullit u mbështet nga Fondacioni Rus për Kërkime Bazë (grantet nr. 99-04-48832; 00-04-49036; 00-04-81086), Federata Ruse për mbështetjen e shkollave shkencore (grantet Nr. 00-115-97833 dhe NSh-1794.2003.4) dhe Programi Akademia Ruse Shkenca “Markerët molekularë gjenetikë dhe kromozomalë në zhvillimin e metodave moderne të mbarështimit dhe prodhimit të farës”.
LITERATURA
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Hyrje në gjenomikën e bimëve // Biologjia molekulare. 2001. V. 35. S. 339-348. 2. Stilolaps E. Bonanza për gjenomikën e bimëve // Shkencë. 1998. V. 282. F. 652-654. 3. Gjenomika e bimëve, Proc. Natl. Akad. shkenca. SHBA. 1998. V. 95. P. 1962-2032. 4. Karteli N.A. dhe etj. Gjenetika. Fjalor Enciklopedik. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferencimi i gjenomit në Aegilops. 1. Shpërndarja e sekuencave të ADN-së shumë të përsëritura në kromozomet e specieve diploide // Gjenomi. 1996. V. 39. F. 293-306. Historia e analizës së kromozomeve // Biol. membranat. 2001. T. 18. S. 164-172.
Më 05/09/2011 në 09:36, Limarev tha:
Limarev V.N.
Deshifrimi i gjenomit njerëzor.
Fragment nga libri i L.G. Puchko: "Njohuri radiologjike e njeriut"
Për zgjidhjen e problemeve të deshifrimit të gjenomit u organizua projekti ndërkombëtar “Gjenomi i njeriut” me një buxhet prej miliarda dollarësh.
Deri në vitin 2000, harta e gjenomit njerëzor ishte praktikisht e plotë. Gjenet u numëruan, u identifikuan dhe u regjistruan në bazat e të dhënave. Këto janë sasi të mëdha informacioni.
Regjistrimi i gjenomit njerëzor në formë të dixhitalizuar kërkon rreth 300 terabajt memorie kompjuterike, që është e barabartë me 3000 disqe të ngurtë me një kapacitet prej 100 gigabajt.
Doli. Se një person nuk ka qindra mijëra, siç mendohej më parë, por pak më shumë se 30 mijë gjene. Miza ka Drosophila, ka vetëm gjysmën e tyre - rreth 13 mijë, dhe miu ka pothuajse të njëjtin numër si një person. Gjenet unike për njerëzit në gjenomin e deshifruar janë vetëm rreth 1%. Pjesa më e madhe e spirales së ADN-së, siç doli, është e zënë jo nga gjenet, por nga të ashtuquajturat "seksione boshe", në të cilat gjenet thjesht nuk janë të koduara, si dhe fragmente të dyfishta që përsëriten njëri pas tjetrit, kuptimi dhe kuptimi. e cila është e paqartë.
Me një fjalë, gjenet rezultuan se nuk ishin as tullat e jetës, por vetëm elemente të planit, sipas të cilit ndërtohet ndërtesa e organizmit. Tullat, siç besohej në gjëra të tjera para lulëzimit të gjenetikës, janë proteina.
U bë absolutisht e qartë se në 1% të gjeneve unike për njerëzit, një sasi kaq e madhe informacioni që i dallon njerëzit nga minjtë nuk mund të kodohet. Ku ruhen të gjitha informacionet? Për shumë shkencëtarë, bëhet një fakt i padyshimtë se pa parimin hyjnor është e pamundur të shpjegohet natyra e njeriut. Një numër shkencëtarësh sugjerojnë se, në kuadrin e ideve ekzistuese për trupin e njeriut, është në thelb e pamundur të deshifrohet gjenomi i njeriut.
Bota nuk njihet - dihet (komentet e mia për artikullin).
1) Shqyrtoni fragmentin: "Pa parimin hyjnor, është e pamundur të shpjegohet natyra e njeriut."
Informacioni i mësipërm nuk thotë asgjë për këtë.
Gjenomi, me të vërtetë, ka një strukturë më komplekse sesa mendohej më parë.
Por, në fund të fundit, kompjuteri i përmendur në artikull nuk përbëhet vetëm nga qelizat e kujtesës.
Një kompjuter ka dy memorie: afatgjatë dhe operacionale, si dhe një procesor në të cilin përpunohet informacioni. Merr pjesë në përpunimin e informacionit dhe fushën elektromagnetike. Për të deshifruar informacionin e gjenomit, është e nevojshme të kuptohet se si ndodh, jo vetëm ruajtja e informacionit, por edhe përpunimi i tij. Unë gjithashtu pranoj idenë se një pjesë e informacionit ruhet e regjistruar nga fushë elektromagnetike. Dhe gjithashtu jashtë një personi, siç kam shkruar tashmë, në qendrat speciale të informacionit të Mendjes së Lartë.
Imagjinoni një tekst të vazhdueshëm të koduar në kodin binar 0 ose 1 të kodit Morse, ndërkohë që nuk dini në çfarë gjuhe (anglisht apo frëngjisht ....) është shkruar dhe nuk e dini se ky tekst i vazhdueshëm përbëhet nga fjalë, fjali. , paragrafë, kapituj, vëllime, rafte, kabinete etj.
Është pothuajse e njëjta gjë në biologji, vetëm këtu gjithçka është e koduar me një kod katërshifror dhe deri më tani kemi deshifruar rendin e gjeneve elementare + - / *, por nuk e dimë gjuhën dhe, në përputhje me rrethanat fjalë, fjali, paragrafë, kapituj, vëllime, rafte, kabinete etj. Gjenomi i deshifruar për ne është ende një tekst i fortë i një kodi me 4 drithëra dhe është pothuajse e pamundur ta studiojmë të gjithë kokë më kokë.
Por rezulton se në periudha të caktuara (si në një individ ashtu edhe në grupin e brezave të tij dhe në një specie, gjini), disa gjene dhe komplekset e tyre (përgjegjëse për fjalët, fjalitë, paragrafët, kapitujt, vëllimet, raftet, kabinetet, etj.). ) janë aktive, dhe në periudha të tjera të evolucionit janë pasive, të cilat i përcaktova në mënyrë indirekte nga tipare të ndryshme poligjenike (që tregohet në temën Ligji Periodik Universal i Evolucionit).
Deri më tani, ekzistojnë vetëm dy metoda për studimin e gjeneve, kjo është një llogaritje e thjeshtë laboratorike e shumës së gjeneve (ADN) në një mostër dhe ekziston një pajisje që numëron sasinë e ARN-së të prodhuar nga proteinat. mbërthyer në çipin elektronik të krijuar ADN specifike, por meqenëse një sasi e madhe e ADN-së është aktive në çdo moment të kohës dhe, në përputhje me rrethanat, një sasi e madhe e proteinave të ndryshme prodhohen përmes ARN-së, është shumë e vështirë të ndash "këtë petë me një lugë, pirun dhe shkopinj japonezë" në këtë supë dhe gjeni atë që kërkoni - të gjeni marrëdhënie shkakësore midis një ADN-je të veçantë (si një kompleks i ADN-së) dhe ndikimit të saj në një tipar poligjenik.
Duket se kam gjetur një metodë të thjeshtë se si të zgjidh të gjithë këtë supë me ADN, ARN dhe proteinat e tyre që përcaktojnë shkallën e një tipari poligjenik.
Siç doli, çdo tipar poligjenik në rendin e evolucionit të një individi (kohorta e brezave, specieve dhe gjinive) është periodike, prandaj, ato duhet të jenë periodike në aktivitetin e ARN-së dhe ADN-së, dhe për këtë arsye është e nevojshme vetëm të gjendet (së pari duke hyrë në detaje gjenetike) një korrelacion midis ndryshimit metrikë në një tipar poligjenik (në një individ, një grup brezash, një specie, një gjini...) dhe proporcional me këto periudha, aktivitetit përkatës të ARN-së, ADN-së.
) gjendet në gjenomën e mizës së frutave ( Drosophila ananassae) një kopje e plotë e gjenomit bakterial të parazitit Wolbachia.
Bakteri Wolbachia jeton në citoplazmën e qelizave pritëse dhe është i njohur për faktin se ka mësuar të rregullojë imtësisht riprodhimin, zhvillimin dhe madje evolucionin e strehuesve të tij. Prandaj, shpesh quhet "manipulues i mikrobeve" ose "zot i mizave" (pasi jeton në qelizat e insekteve).
Studimi filloi kur Julie Dunning-Hotopp e JCVI zbuloi se si disa gjene Wolbachia "bashkëpunojnë" me gjenet e Drosophila sikur të ishin pjesë e të njëjtit gjenom.
Michael Clark - studiues në Universitetin e Rochester - vendosi koloninë Drosophila ananassae në laborator për të zbuluar sekretin me Warren.
Gjeni Wolbachia në gjenomën Drosophila (ilustruar nga Universiteti i Rochester).
"Për disa muaj, mendova se e kisha gabim për diçka," thotë Clarke, "madje sugjerova se ishte zhvilluar rezistenca ndaj antibiotikëve, sepse gjeta çdo gjen Wolbachia pa pushim. Kur më në fund mora shamitë që kisha lënë vetëm disa muaj më parë, nuk e gjeta vetë Wolbachia.
Tani Warren dhe Clark po përpiqen të kuptojnë se cili është avantazhi i futjes së një pjese kaq të madhe të ADN-së për Drosophila - ndoshta gjenet "aliene" i ofrojnë hostit disa mundësi të reja.

Dhe kështu gjenet Wolbachia kalojnë në ADN-në e pritësit (ilustrim nga Nicolle Rager Fuller, National Science).
Rezultatet e studimit u publikuan në një artikull në revistën Science. Në të, autorët sugjerojnë se transferimi horizontal i gjeneve (transferimi i gjeneve midis specieve të palidhura) ndodh midis baktereve dhe organizmave shumëqelizorë në botën tonë shumë më shpesh sesa mendohej më parë.
Deshifrimi i mekanizmave gjenetikë molekularë të manipulimeve të kryera nga Wolbachia me nikoqirët e saj do t'i japë njeriut mjete të reja të fuqishme për të ndikuar në organizmat e gjallë dhe natyrën në tërësi.
Sidoqoftë, jo të gjitha insektet i nënshtrohen ndikimit të keq të Wolbachia. Për shembull, fluturat nga Ishujt Samoan kanë "mësuar" të mbrojnë meshkujt e tyre. Pyes veten nëse mushkonjat e malaries, të cilat duan t'i infektojnë me këtë bakter, do të mësojnë ta luftojnë atë?
Për 50 vjetorin e zbulimit të strukturës së ADN-së
A.V. Zelenin
GJENOMI I BIMËVE
A. V. Zelenin
Zelenin Alexander Vladimirovich- d.b.n.,
shef i laboratorit të Institutit të Biologjisë Molekulare. V.A. Engelhardt RAS.
Arritjet mbresëlënëse të programit "Genomi i njeriut", si dhe suksesi i punës për deshifrimin e të ashtuquajturave gjenome ekstra të vogla (viruse), të vogla (baktere, maja) dhe të mesme (krimbët e rrumbullakët, Drosophila), bënë të mundur që të kaloni në një studim në shkallë të gjerë të gjenomave të bimëve të mëdha dhe tepër të mëdha. Nevoja urgjente për një studim të hollësishëm të gjenomave të bimëve më të rëndësishme ekonomikisht u theksua në një takim mbi gjenomikën e bimëve të mbajtur në vitin 1997 në Shtetet e Bashkuara [ , ]. Gjatë viteve që kanë kaluar që nga ajo kohë, janë arritur suksese të padyshimta në këtë fushë. Në vitin 2000, u shfaq një botim mbi sekuencën e plotë (përcaktimin e sekuencës nukleotide lineare të të gjithë ADN-së bërthamore) të gjenomit të mustardës së vogël - Arabidopsis, në 2001 - mbi sekuencën paraprake (draft) të gjenomit të orizit. Punimet për renditjen e gjenomave të bimëve të mëdha dhe super të mëdha (misër, thekër, grurë) u raportuan vazhdimisht, megjithatë, këto raporte nuk përmbanin informacion specifik dhe ishin më tepër në natyrën e deklaratave të qëllimit.
Supozohet se dekodimi i gjenomave të bimëve do të hapë perspektiva të gjera për shkencën dhe praktikën. Para së gjithash, identifikimi i gjeneve të reja dhe zinxhiri i rregullimit të tyre gjenetik do të rrisë ndjeshëm produktivitetin e bimëve nëpërmjet përdorimit të qasjeve bioteknologjike. Me zbulimin, izolimin, riprodhimin (klonimin) dhe renditjen e gjeneve përgjegjëse për funksione kaq të rëndësishme të organizmit bimor si riprodhimi dhe produktiviteti, proceset e ndryshueshmërisë, rezistenca ndaj faktorëve të pafavorshëm mjedisor, si dhe çiftimi homolog i kromozomeve, shfaqja e shoqërohen mundësi të reja për përmirësimin e procesit të mbarështimit. Së fundi, gjenet e izoluara dhe të klonuara mund të përdoren për të marrë bimë transgjenike me veti thelbësisht të reja dhe për të analizuar mekanizmat e rregullimit të aktivitetit të gjeneve.
Rëndësia e studimit të gjenomave të bimëve theksohet edhe nga fakti se deri tani numri i gjeneve të bimëve të lokalizuara, të klonuara dhe të sekuencave është i vogël dhe varion, sipas vlerësimeve të ndryshme, midis 800 dhe 1200. Kjo është 10-15 herë më pak se, për për shembull, te njerëzit.
Shtetet e Bashkuara mbeten lideri i padyshimtë në studimin në shkallë të gjerë të gjenomave të bimëve, megjithëse studime intensive të gjenomit të orizit po kryhen në Japoni dhe, vitet e fundit, në Kinë. Në deshifrimin e gjenomit të Arabidopsis, përveç laboratorëve amerikanë, morën pjesë aktive edhe grupe kërkimore evropiane. Udhëheqja e dukshme e Shteteve të Bashkuara shkakton shqetësim serioz të shkencëtarëve evropianë, të cilin ata e shprehën qartë në një takim me titullin domethënës "Perspektivat për gjenomikën në epokën post-gjenomike", mbajtur në fund të vitit 2000 në Francë. Përparimi i shkencës amerikane në studimin e gjenomit të bimëve bujqësore dhe krijimin e formave të bimëve transgjenike, sipas shkencëtarëve evropianë, kërcënon se në një të ardhme jo shumë të largët (dy deri në pesë dekada), kur rritja e popullsisë do ta vendosë njerëzimin përballë një kriza ushqimore, ekonomia dhe shkenca evropiane do të varen nga teknologjia amerikane. Në këtë drejtim u njoftua krijimi i një programi shkencor franko-gjerman për studimin e gjenomave të bimëve (“Plantgene”) dhe në të u bënë investime të konsiderueshme.
Natyrisht, problemet e gjenomikës së bimëve duhet të tërheqin vëmendjen e ngushtë të shkencëtarëve dhe organizatorëve rusë të shkencës, si dhe të organeve drejtuese, pasi nuk ka të bëjë vetëm me prestigjin shkencor, por edhe me sigurinë kombëtare të vendit. Në një ose dy dekada, ushqimi do të bëhet burimi më i rëndësishëm strategjik.
VËSHTIRËSITË NË STUDIMIN E GJENOMËVE TË BIMËVE
Studimi i gjenomit të bimëve është një detyrë shumë më e vështirë sesa studimi i gjenomit të njerëzve dhe kafshëve të tjera. Kjo është për shkak të rrethanave të mëposhtme:
madhësi të mëdha gjenomi, që arrijnë dhjetëra dhe madje qindra miliarda çifte bazë (bp) për speciet individuale bimore: gjenomet e bimëve kryesore ekonomikisht të rëndësishme (përveç orizit, lirit dhe pambukut) janë ose të përmasave afër gjenomit njerëzor, ose e tejkaloni atë shumë herë (tabela);STUDIME KROMOZOMALE TË GJENOMEVELuhatje të mprehta në numrin e kromozomeve në bimë të ndryshme - nga dy në disa specie në disa qindra në të tjera, dhe nuk është e mundur të identifikohet një korrelacion i rreptë midis madhësisë së gjenomit dhe numrit të kromozomeve;
Një bollëk poliploidësh (që përmbajnë më shumë se dy gjenome për qelizë) me gjenome të ngjashme, por jo identike (alpolyploidy);
Pasurimi ekstrem i gjenomave të bimëve (deri në 99%) të ADN-së "të parëndësishme" (jo koduese, d.m.th., që nuk përmban gjene), e cila e ndërlikon shumë ngjitjen (rregullimin në rendin e duhur) të fragmenteve të sekuencave në një të përbashkët të gjerë. rajoni i ADN-së me madhësi (contig);
Harta morfologjike, gjenetike dhe fizike e kromozomeve jo të plota (në krahasim me gjenomin Drosophila, të njeriut dhe të miut);
Pamundësia praktike e izolimit të kromozomeve individuale në formë të pastër duke përdorur metoda që zakonisht përdoren për këtë qëllim për kromozomet e njeriut dhe të kafshëve (ndarja në një rrjedhë dhe përdorimi i hibrideve qelizore);
Vështirësia e hartës kromozomike (përcaktimi i vendndodhjes në kromozom) të gjeneve individuale duke përdorur hibridizimin në vend, si për shkak të përmbajtjes së lartë të ADN-së "të parëndësishme" në gjenomet e bimëve, ashtu edhe për veçoritë e organizimit strukturor të kromozomeve bimore;
Largësia evolucionare e bimëve nga kafshët, e cila ndërlikon seriozisht përdorimin e informacionit të marrë nga renditja e gjenomave të njerëzve dhe kafshëve të tjera për studimin e gjenomave të bimëve;
Procesi i gjatë i riprodhimit të shumicës së bimëve, i cili ngadalëson ndjeshëm analizën e tyre gjenetike.
Studimet kromozomale (citogjenetike) të gjenomave në përgjithësi dhe bimëve në veçanti kanë një histori të gjatë. Termi "gjenom" u propozua për të treguar një grup haploid (të vetëm) kromozomesh me gjenet e përfshira në to në çerekun e parë të shekullit të 20-të, domethënë shumë kohë përpara vendosjes së rolit të ADN-së si bartës i informacionit gjenetik. .
Përshkrimi i gjenomit të një organizmi shumëqelizor të ri, të pastudiuar gjenetikisht më parë, zakonisht fillon me studimin dhe përshkrimin e grupit të plotë të kromozomeve të tij (kariotip). Kjo, natyrisht, vlen edhe për bimët, një numër i madh i të cilave as nuk kanë filluar të studiohen.
Tashmë në agimin e studimeve të kromozomeve, gjenomet e specieve bimore të lidhura u krahasuan bazuar në analizën e konjugimit mejotik (kombinimi i kromozomeve homologe) në hibridet ndërspecifike. Gjatë 100 viteve të fundit, mundësitë e analizës së kromozomeve janë zgjeruar në mënyrë dramatike. Tani, teknologjitë më të avancuara përdoren për të karakterizuar gjenomet e bimëve: variante të ndryshme të të ashtuquajturës ngjyrosje diferenciale, e cila bën të mundur identifikimin e kromozomeve individuale sipas veçorive morfologjike; hibridizimi në vend duke bërë të mundur lokalizimin e gjeneve specifike në kromozome; studimet biokimike të proteinave qelizore (elektroforeza dhe imunokimia) dhe, së fundi, një grup metodash të bazuara në analizën e ADN-së kromozomale deri në renditjen e saj.

Oriz. një. Kariotipet e drithërave a - thekra (14 kromozome), b - gruri i fortë (28 kromozome), c - gruri i butë (42 kromozome), d - elbi (14 kromozome)Për shumë vite, janë studiuar kariotipet e drithërave, kryesisht gruri dhe thekra. Është interesante se në lloje të ndryshme të këtyre bimëve, numri i kromozomeve është i ndryshëm, por gjithmonë shumëfish i shtatë. Llojet individuale të drithërave mund të njihen me besueshmëri nga kariotipi i tyre. Për shembull, gjenomi i thekrës përbëhet nga shtatë çifte kromozomesh të mëdhenj me blloqe heterokromatike me ngjyra intensive në skajet e tyre, shpesh të quajtura segmente ose breza (Fig. 1a). Gjenomet e grurit kanë tashmë 14 dhe 21 çifte kromozomesh (Fig. 1, b, c), dhe shpërndarja e blloqeve heterokromatike në to nuk është e njëjtë si në kromozomet e thekrës. Gjenomet individuale të grurit, të emërtuara A, B dhe D, gjithashtu ndryshojnë nga njëra-tjetra. Një rritje në numrin e kromozomeve nga 14 në 21 çon në një ndryshim të mprehtë në vetitë e grurit, gjë që pasqyrohet në emrat e tyre: durum, ose makarona, grurë dhe të buta, ose bukë, grurë . Gjeni D, i cili përmban gjene për proteinat e glutenit, i cili i jep brumit të ashtuquajturin mbirje, është përgjegjës për përvetësimin e vetive të larta të pjekjes nga gruri i butë. Pikërisht këtij gjenomi i kushtohet vëmendje e veçantë në përmirësimin e përzgjedhjes së grurit të bukës. Një tjetër drithëra me 14 kromozome, elbi (Fig. 1, d), zakonisht nuk përdoret për të bërë bukë, por është lënda e parë kryesore për prodhimin e produkteve të tilla të zakonshme si birra dhe uiski.
Po studiohen intensivisht kromozomet e disa bimëve të egra që përdoren për të përmirësuar cilësinë e specieve më të rëndësishme bujqësore, siç janë të afërmit e egër të grurit - Aegilops. Format e reja bimore krijohen nga kryqëzimi (Fig. 2) dhe seleksionimi. Vitet e fundit, një përmirësim i ndjeshëm i metodave të kërkimit ka bërë të mundur fillimin e studimit të gjenomave të bimëve, tiparet e kariotipeve të të cilave (kryesisht madhësia e vogël e kromozomeve) i bënë ato më parë të paarritshme për analizën e kromozomeve. Pra, vetëm kohët e fundit të gjitha kromozomet e pambukut, kamomilit dhe lirit u identifikuan për herë të parë.

Oriz. 2. Kariotipet e grurit dhe një hibrid i grurit me Aegilops
a - grurë i butë heksaploid ( Triticum astivum), i përbërë nga gjenomet A, B dhe O; b - grurë tetraploid ( Triticum timopheevi), i përbërë nga gjenomet A dhe G. përmban gjene për rezistencë ndaj shumicës së sëmundjeve të grurit; c - hibride Triticum astivum X Triticum timopheevi rezistent ndaj mykut pluhur dhe ndryshkut, zëvendësimi i një pjese të kromozomeve është qartë i dukshëmSTRUKTURA PRIMARE E ADN-së
Me zhvillimin e gjenetikës molekulare, vetë koncepti i gjenomit është zgjeruar. Tani ky term interpretohet si në kromozomin klasik ashtu edhe në kuptimin molekular modern: i gjithë materiali gjenetik i një virusi, qelize dhe organizmi individual. Natyrisht, pas studimit të strukturës së plotë parësore të gjenomave (siç quhet shpesh sekuenca e plotë lineare e bazave të acidit nukleik) të një numri mikroorganizmash dhe njerëzve, u ngrit çështja e renditjes së gjenomit bimor.
Nga shumë organizma bimorë, dy u zgjodhën për studim - Arabidopsis, që përfaqëson klasën e dikoteve (madhësia e gjenomit 125 milion bp) dhe orizi nga klasa e monocots (420-470 milion bp). Këto gjenome janë të vogla në krahasim me gjenomet e tjera bimore dhe përmbajnë relativisht pak segmente të përsëritura të ADN-së. Veçori të tilla dhanë shpresë se gjenomet e përzgjedhura do të ishin të disponueshme për përcaktimin relativisht të shpejtë të strukturës së tyre parësore.

Oriz. 3. Arabidopsis - mustardë e vogël - një bimë e vogël nga familja e kryqëzatave ( Brassicaceae). Në një hapësirë të barabartë në sipërfaqe me një faqe të revistës sonë, ju mund të rritni deri në një mijë organizma individualë Arabidopsis.Arsyeja e zgjedhjes së Arabidopsis nuk ishte vetëm madhësia e vogël e gjenomit të saj, por edhe madhësia e vogël e organizmit, gjë që e bën të lehtë rritjen e saj në laborator (Fig. 3). Ne morëm parasysh ciklin e tij të shkurtër riprodhues, falë të cilit është e mundur të kryhen shpejt eksperimente për kryqëzimin dhe përzgjedhjen, gjenetikën e studiuar në detaje, lehtësinë e manipulimit me ndryshimin e kushteve të rritjes (ndryshimi i përbërjes së kripës së tokës, shtimi i lëndëve ushqyese të ndryshme, etj. .) dhe testimi i efektit në bimë të faktorëve dhe patogjenëve të ndryshëm mutagjenë (viruse, baktere, kërpudha). Arabidopsis nuk ka asnjë vlerë ekonomike, prandaj gjenomi i tij, së bashku me gjenomin e miut, u quajt referencë, ose, më pak saktë, model.*
* Shfaqja e termit "gjenom model" në literaturën ruse është rezultat i një përkthimi të pasaktë të frazës angleze model genome. Fjala "model" nënkupton jo vetëm mbiemrin "model", por edhe emrin "kampion", "standard", "model". Do të ishte më e saktë të flitej për një gjenom mostër, ose një gjenom referencë.Puna intensive për renditjen e gjenomit të Arabidopsis filloi në vitin 1996 nga një konsorcium ndërkombëtar që përfshinte institucione shkencore dhe grupe kërkimore nga SHBA, Japonia, Belgjika, Italia, Britania e Madhe dhe Gjermania. Në dhjetor 2000, informacione të gjera u vunë në dispozicion duke përmbledhur përcaktimin e strukturës parësore të gjenomit Arabidopsis. Për renditjen u përdor teknologjia klasike ose hierarkike: së pari, u studiuan seksione të vogla individuale të gjenomit, nga të cilat përbëheshin seksione më të mëdha (contigs) dhe, në fazën përfundimtare, struktura e kromozomeve individuale. ADN-ja bërthamore e gjenomit Arabidopsis shpërndahet në pesë kromozome. Në vitin 1999, u publikuan rezultatet e sekuencës së dy kromozomeve dhe shfaqja në shtyp e informacionit në lidhje me strukturën parësore të tre të tjerëve përfundoi sekuencën e të gjithë gjenomit.
Nga 125 milionë çifte bazash, është përcaktuar struktura primare prej 119 milionë, që është 92% e të gjithë gjenomit. Vetëm 8% e gjenomit Arabidopsis që përmban blloqe të mëdha të segmenteve të përsëritura të ADN-së doli të ishte i paarritshëm për studim. Për sa i përket tërësisë dhe plotësimit të sekuencës së gjenomit eukariotik, Arabidopsis mbetet në tre kampionët kryesorë së bashku me një organizëm majaje njëqelizore. Saccharomyces cerevisiae dhe organizmit shumëqelizor Eleganca e Caenorhabditis(shih tabelën).
Rreth 15,000 gjene individuale të kodimit të proteinave janë gjetur në gjenomën Arabidopsis. Përafërsisht 12,000 prej tyre përmbahen si dy kopje për gjenom haploid (të vetëm), kështu që numri i përgjithshëm i gjeneve është 27,000. Numri i gjeneve në Arabidopsis nuk ndryshon shumë nga numri i gjeneve në organizmat si njerëzit dhe minjtë, por madhësia e gjenomit të tij 25-30 herë më pak. Kjo rrethanë lidhet me karakteristika të rëndësishme në strukturën e gjeneve individuale të Arabidopsis dhe strukturën e përgjithshme të gjenomit të tij.
Gjenet e Arabidopsis janë kompakte, që përmbajnë vetëm disa ekzone (rajone koduese të proteinave) të ndara nga segmente të shkurtra (rreth 250 bp) jo-koduese të ADN-së (introne). Intervalet ndërmjet gjeneve individuale janë mesatarisht 4600 çifte bazash. Për krahasim, theksojmë se gjenet njerëzore përmbajnë shumë dhjetëra dhe madje qindra ekzone dhe introne, dhe rajonet ndërgjenike kanë madhësi prej 10 mijë çifte bazash ose më shumë. Supozohet se prania e një gjenomi të vogël kompakt kontribuoi në stabilitetin evolucionar të Arabidopsis, pasi ADN-ja e tij u bë një objektiv për agjentë të ndryshëm dëmtues në një masë më të vogël, në veçanti, për futjen e fragmenteve të ADN-së të ngjashme me virusin (transpozonët). në gjenom.
Ndër veçoritë e tjera molekulare të gjenomit Arabidopsis, duhet theksuar se ekzonet pasurohen me guaninë dhe citozinë (44% në ekzone dhe 32% në introne) krahasuar me gjenet e kafshëve, si dhe prania e gjeneve të përsëritura dyfish (të dyfishuara). Supozohet se një dyfishim i tillë ka ndodhur si rezultat i katër ngjarjeve të njëkohshme, që konsistojnë në dyfishimin (përsëritjen) e një pjese të gjeneve Arabidopsis, ose shkrirjen e gjenomave të lidhura me to. Këto ngjarje, të ndodhura 100-200 milion vjet më parë, janë një manifestim i prirjes së përgjithshme drejt poliploidizimit (rritje e shumëfishtë e numrit të gjenomave në një organizëm), që është karakteristikë e gjenomave të bimëve. Megjithatë, disa fakte tregojnë se gjenet e dyfishta në Arabidopsis nuk janë identike dhe funksionojnë ndryshe, gjë që mund të shoqërohet me mutacione në rajonet e tyre rregullatore.
Orizi është bërë një tjetër objekt i sekuencës së plotë të ADN-së. Gjenomi i kësaj bime është gjithashtu i vogël (12 kromozome, që japin një total prej 420-470 milion bp), vetëm 3,5 herë më i madh se ai i Arabidopsis. Sidoqoftë, ndryshe nga Arabidopsis, orizi ka një rëndësi të madhe ekonomike, duke qenë baza e të ushqyerit për më shumë se gjysmën e njerëzimit, prandaj, jo vetëm miliarda konsumatorë, por edhe një ushtri shumëmilionëshe njerëzish të përfshirë në mënyrë aktive në procesin shumë të mundimshëm të tij. kultivimi janë jashtëzakonisht të interesuar në përmirësimin e vetive të tij.
Disa studiues filluan të studiojnë gjenomën e orizit që në vitet 1980, por këto studime arritën një shkallë serioze vetëm në vitet 1990. Në vitin 1991, në Japoni u krijua një program për të deshifruar strukturën e gjenomit të orizit, duke bashkuar përpjekjet e shumë grupeve kërkimore. Në vitin 1997, mbi bazën e këtij programi u organizua Projekti Ndërkombëtar i Gjenomit të Orizit. Pjesëmarrësit e tij vendosën të përqendrojnë përpjekjet e tyre në renditjen e një prej nënspecieve të orizit ( Oriza sativajaponica), në studimin e së cilës ishte arritur tashmë përparim i rëndësishëm deri në atë kohë. Një stimul serioz dhe, në mënyrë figurative, një yll udhëzues për një punë të tillë ishte programi "Gjenomi i njeriut".
Në kuadër të këtij programi u testua strategjia e ndarjes hierarkike “kromozomale” të gjenomit, të cilën pjesëmarrësit e konsorciumit ndërkombëtar e përdorën për deshifrimin e gjenomit të orizit. Sidoqoftë, nëse në studimin e gjenomit njerëzor, fraksionet e kromozomeve individuale u izoluan duke përdorur metoda të ndryshme, atëherë materiali specifik për kromozomet individuale të orizit dhe rajonet e tyre individuale u mor me mikrodiseksion lazer (prerja e objekteve mikroskopike). Në një rrëshqitje mikroskopi, ku ndodhen kromozomet e orizit, nën ndikimin e një rreze lazer, gjithçka digjet, përveç kromozomit ose pjesëve të tij të planifikuara për analizë. Materiali i mbetur përdoret për klonim dhe sekuencë.
Janë publikuar raporte të shumta mbi rezultatet e renditjes së fragmenteve individuale të gjenomës së orizit, të kryera me saktësi dhe detaje të larta, karakteristikë e teknologjisë hierarkike. Besohej se përcaktimi i strukturës së plotë parësore të gjenomit të orizit do të përfundonte në fund të vitit 2003-mesi i vitit 2004 dhe rezultatet, së bashku me të dhënat mbi strukturën parësore të gjenomit Arabidopsis, do të përdoreshin gjerësisht në krahasimin. gjenomika e bimëve të tjera.
Megjithatë, në fillim të vitit 2002, dy grupe kërkimore - njëri nga Kina, tjetri nga Zvicra dhe Shtetet e Bashkuara - publikuan rezultatet e një drafti të plotë (të përafërt) të sekuencës së gjenomit të orizit, të kryer duke përdorur teknologjinë e klonimit total. Në ndryshim nga studimi i staduar (hierarkik), qasja totale bazohet në klonimin e njëkohshëm të të gjithë ADN-së gjenomike në një nga vektorët viralë ose bakterialë dhe marrjen e një numri të konsiderueshëm (të madh për gjenomet e mesme dhe të mëdha) të kloneve individuale që përmbajnë të ndryshëm Segmentet e ADN-së. Bazuar në analizën e këtyre seksioneve të sekuencave dhe mbivendosjes së seksioneve identike terminale të ADN-së, formohet një kontig - një zinxhir sekuencash të ADN-së të bashkuara së bashku. Kontigu i përgjithshëm (gjithsej) është struktura primare e të gjithë gjenomit, ose të paktën e një kromozomi individual.
Në një paraqitje të tillë skematike, strategjia e klonimit total duket e thjeshtë. Në fakt, ai ndeshet me vështirësi serioze që lidhen me nevojën për të marrë një numër të madh klonesh (përgjithësisht pranohet që gjenomi ose rajoni i tij në studim duhet të mbivendosen nga klone të paktën 10 herë), sasinë e madhe të sekuencave dhe jashtëzakonisht punë komplekse e kloneve docking që kërkojnë pjesëmarrjen e specialistëve të bioinformatikës. Një pengesë serioze për klonimin total është një shumëllojshmëri segmentesh të përsëritura të ADN-së, numri i të cilave, siç u përmend tashmë, rritet ndjeshëm me rritjen e madhësisë së gjenomit. Prandaj, strategjia e sekuencës totale përdoret kryesisht në studimin e gjenomave të viruseve dhe mikroorganizmave, megjithëse është përdorur me sukses për të studiuar gjenomin e një organizmi shumëqelizor, Drosophila.
Rezultatet e sekuencës totale të këtij gjenomi u "mbivendosën" në një grup të madh informacioni rreth strukturës së tij kromozomale, gjenesh dhe molekulare të marra gjatë një periudhe pothuajse 100-vjeçare të studimit të Drosophila. E megjithatë, për sa i përket shkallës së renditjes, gjenomi Drosophila (66% e madhësisë totale të gjenomit) është dukshëm inferior ndaj gjenomit të Arabidopsis (92%), megjithë madhësitë e tyre mjaft të afërta - përkatësisht 180 milion dhe 125 milion çifte bazë. . Prandaj, kohët e fundit është propozuar të emërtohet teknologjia e përzier, e cila u përdor për sekuencimin e gjenomit Drosophila.
Për të renditur gjenomin e orizit, grupet kërkimore të përmendura më sipër morën dy nga nënllojet e tij, më të kultivuarat në vendet aziatike, - Oriza saliva L. ssp indicaj dhe Oriza saliva L. sspjaponica. Rezultatet e studimeve të tyre përkojnë në shumë aspekte, por ndryshojnë në shumë aspekte. Kështu, përfaqësuesit e të dy grupeve deklaruan se kishin arritur afërsisht 92-93% të mbivendosjes së gjenomit me kontigs. Është treguar se rreth 42% e gjenomës së orizit përfaqësohet nga përsëritje të shkurtra të ADN-së që përbëhen nga 20 çifte bazash, dhe shumica e elementëve të lëvizshëm të ADN-së (transpozonëve) ndodhen në rajone ndërgjenike. Megjithatë, të dhënat për madhësinë e gjenomit të orizit ndryshojnë ndjeshëm.
Për nëngrupin japonez, madhësia e gjenomit është përcaktuar të jetë 466 milionë çifte bazë, dhe për nëngrupin indian, 420 milionë. Arsyeja e kësaj mospërputhjeje nuk është e qartë. Mund të jetë pasojë e qasjeve të ndryshme metodologjike në përcaktimin e madhësisë së pjesës jokoduese të gjenomit, domethënë nuk pasqyron gjendjen e vërtetë të punëve. Por është e mundur që një ndryshim prej 15% në madhësinë e gjenomave të studiuara ekziston.
Mospërputhja e dytë e madhe u zbulua në numrin e gjeneve të gjetura: për nëngrupin japonez, nga 46,022 në 55,615 gjene për gjenom, dhe për nënllojet indiane, nga 32,000 në 50,000. Arsyeja për këtë mospërputhje nuk është e qartë.
Paplotësia dhe mospërputhja e informacionit të marrë vërehet në komentet e artikujve të botuar. Shpresa shprehet gjithashtu këtu se boshllëqet në njohuritë e gjenomit të orizit do të eliminohen duke krahasuar të dhënat e "sekuencës së përafërt" me rezultatet e renditjes së detajuar hierarkike të kryer nga pjesëmarrësit e Projektit Ndërkombëtar të Gjenomit të Orizit.
GJENOMIA KRAHASUESE DHE FUNKSIONALE E BIMËVE
Të dhënat e gjera të marra, gjysma e të cilave (rezultatet e grupit kinez) janë të disponueshme publikisht, hapin padyshim perspektiva të gjera si për studimin e gjenomit të orizit ashtu edhe për gjenomikën e bimëve në përgjithësi. Krahasimi i vetive të gjenomit të Arabidopsis dhe orizit tregoi se shumica e gjeneve (deri në 80%) të identifikuara në gjenomin Arabidopsis gjenden gjithashtu në gjenomën e orizit, megjithatë, për afërsisht gjysmën e gjeneve të gjetura në oriz, analoge (ortologë ) nuk janë gjetur ende në gjenomin Arabidopsis. . Në të njëjtën kohë, 98% e gjeneve, struktura primare e të cilave është krijuar për drithëra të tjera, u gjetën në gjenomën e orizit.
Mospërputhja e rëndësishme (pothuajse e dyfishtë) midis numrit të gjeneve në oriz dhe Arabidopsis është e çuditshme. Në të njëjtën kohë, të dhënat e draftit të dekodimit të gjenomës së orizit, të marra duke përdorur sekuencën totale, praktikisht nuk krahasohen me rezultatet e gjera të studimit të gjenomit të orizit me metodën e klonimit dhe sekuencës hierarkike, domethënë atë që ka është bërë në lidhje me gjenomin Drosophila nuk është kryer. Prandaj, mbetet e paqartë nëse ndryshimi në numrin e gjeneve në Arabidopsis dhe oriz pasqyron gjendjen e vërtetë të punëve apo nëse shpjegohet me ndryshimin në qasjet metodologjike.
Në ndryshim nga gjenomi i Arabidopsis, të dhënat për gjenet binjake në gjenomën e orizit nuk janë dhënë. Është e mundur që sasia e tyre relative të jetë më e lartë në oriz sesa në Arabidopsis. Kjo mundësi mbështetet në mënyrë indirekte nga të dhënat për praninë e formave poliploide të orizit. Më shumë qartësi për këtë çështje mund të pritet pasi të përfundojë Projekti Ndërkombëtar i Gjenomit të Orizit dhe të merret një pamje e detajuar e strukturës primare të ADN-së të këtij gjenomi. Baza serioze për një shpresë të tillë jep fakti se pas botimit të punimeve mbi sekuencën e përafërt të gjenomit të orizit, numri i botimeve mbi strukturën e këtij gjenomi është rritur ndjeshëm, në veçanti, informacioni është shfaqur për renditjen e detajuar. të kromozomeve 1 dhe 4 të tij.
Njohja, të paktën përafërsisht, e numrit të gjeneve në bimë është me rëndësi themelore për gjenomikën krahasuese të bimëve. Fillimisht, besohej se duke qenë se të gjitha bimët e lulëzuara janë shumë afër njëra-tjetrës për sa i përket karakteristikave të tyre fenotipike, gjenomet e tyre gjithashtu duhet të jenë të ngjashme. Dhe nëse studiojmë gjenomin e Arabidopsis, do të marrim informacion për shumicën e gjenomave të bimëve të tjera. Një konfirmim indirekt i këtij supozimi janë rezultatet e sekuencës së gjenomit të miut, i cili është çuditërisht afër gjenomit njerëzor (rreth 30 mijë gjene, nga të cilat vetëm 1 mijë rezultuan të jenë të ndryshëm).
Mund të supozohet se arsyeja e ndryshimeve midis gjenomave të Arabidopsis dhe orizit qëndron në përkatësinë e tyre ndaj klasave të ndryshme të bimëve - dikot dhe monokotë. Për të sqaruar këtë çështje, është shumë e dëshirueshme të njihni të paktën një strukturë primare të përafërt të ndonjë bime tjetër monokotiledone. Kandidati më real mund të jetë misri, gjenomi i të cilit është afërsisht i barabartë me gjenomin e njeriut, por gjithsesi shumë më i vogël se gjenomi i drithërave të tjera. Vlera ushqyese e misrit është e njohur mirë.
Materiali i madh i marrë si rezultat i sekuencës së gjenomit të Arabidopsis dhe orizit po bëhet gradualisht baza për një studim në shkallë të gjerë të gjenomave të bimëve duke përdorur gjenomikën krahasuese. Studime të tilla kanë një rëndësi të përgjithshme biologjike, pasi ato bëjnë të mundur përcaktimin e parimeve kryesore të organizimit të gjenomit të bimëve në tërësi dhe kromozomeve të tyre individuale, për të identifikuar tiparet e përbashkëta të strukturës së gjeneve dhe rajoneve të tyre rregullatore, dhe për të marrë në konsideratë raporti i pjesës (gjenit) funksionalisht aktive të kromozomit dhe rajoneve të ndryshme ndërgjenike të ADN-së që nuk kodojnë për proteinat. Gjenetika krahasuese po bëhet gjithashtu gjithnjë e më e rëndësishme për zhvillimin e gjenomikës funksionale njerëzore. Pikërisht për studime krahasuese u krye sekuenca e gjenomit të peshkut puffer dhe të miut.
Po aq i rëndësishëm është studimi i gjeneve individuale përgjegjëse për sintezën e proteinave individuale që përcaktojnë funksione specifike të trupit. Pikërisht në zbulimin, izolimin, renditjen dhe përcaktimin e funksionit të gjeneve individuale qëndron rëndësia praktike, kryesisht mjekësore, e programit të Gjenomit Njerëzor. Kjo rrethanë u vu në dukje disa vite më parë nga J. Watson, i cili theksoi se programi "Gjenomi i njeriut" do të përfundonte vetëm kur të përcaktoheshin funksionet e të gjitha gjeneve njerëzore.
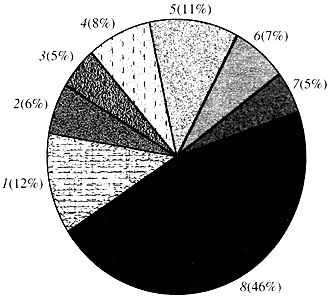
Oriz. katër. Klasifikimi sipas funksionit të gjeneve të Arabidopsis
1 - gjenet për rritjen, ndarjen dhe sintezën e ADN-së; 2 - gjenet e sintezës së ARN-së (transkriptimi); 3 - gjenet për sintezën dhe modifikimin e proteinave; 4 - gjenet për zhvillimin, plakjen dhe vdekjen e qelizave; 5 - gjenet e metabolizmit qelizor dhe metabolizmit të energjisë; 6 - gjenet e ndërveprimit ndërqelizor dhe transmetimit të sinjalit; 7 - gjenet për sigurimin e proceseve të tjera qelizore; 8 - gjene me funksion të panjohurSa i përket funksionit të gjeneve të bimëve, ne dimë më pak se një të dhjetën e asaj që dimë për gjenet njerëzore. Edhe në Arabidopsis, gjenomi i të cilit është shumë më i studiuar se gjenomi i njeriut, funksioni i pothuajse gjysmës së gjeneve të tij mbetet i panjohur (Fig. 4). Ndërkohë, përveç gjeneve të përbashkëta me kafshët, bimët kanë një numër të konsiderueshëm gjenesh që janë specifike vetëm (ose të paktën kryesisht) për to. Bëhet fjalë për gjenet që marrin pjesë në transportin e ujit dhe sintezën e murit qelizor, i cili mungon tek kafshët, gjenet që sigurojnë formimin dhe funksionimin e kloroplasteve, fotosintezën, fiksimin e azotit dhe sintezën e produkteve të shumta aromatike. Kjo listë mund të vazhdojë, por tashmë është e qartë se çfarë detyre të vështirë përballet gjenomika funksionale e bimëve.
Sekuenca e plotë e gjenomit siguron informacion afërsisht të vërtetë për numrin total të gjeneve në një organizëm të caktuar, bën të mundur vendosjen e informacionit pak a shumë të detajuar dhe të besueshëm për strukturën e tyre në bankat e të dhënave dhe lehtëson punën e izolimit dhe studimit të gjeneve individuale. Megjithatë, sekuenca e gjenomit nuk do të thotë në asnjë mënyrë vendosjen e funksionit të të gjitha gjeneve.
Një nga qasjet më premtuese të gjenomikës funksionale bazohet në identifikimin e gjeneve të punës që përdoren për transkriptimin (leximin) e mARN-së. Kjo qasje, duke përfshirë përdorimin e teknologjisë moderne të mikrogrumeve, bën të mundur identifikimin e njëkohshëm të deri në dhjetëra mijëra gjeneve funksionale. Kohët e fundit, duke përdorur këtë qasje, ka filluar studimi i gjenomave të bimëve. Për Arabidopsis, ishte e mundur të merreshin rreth 26 mijë transkripte individuale, gjë që lehtëson shumë mundësinë e përcaktimit të funksionit të pothuajse të gjithë gjeneve të tij. Në patate, ishte e mundur të identifikoheshin rreth 20,000 gjene funksionale që janë të rëndësishme për të kuptuar si proceset e rritjes dhe formimit të tuberit, ashtu edhe proceset e sëmundjes së patates. Supozohet se kjo njohuri do të rrisë rezistencën e një prej produkteve ushqimore më të rëndësishme ndaj patogjenëve.
Zhvillimi logjik i gjenomikës funksionale ishte proteomika. Kjo fushë e re e shkencës studion proteomën, e cila zakonisht kuptohet si grupi i plotë i proteinave në një qelizë në një moment të caktuar. Një grup i tillë proteinash, duke pasqyruar gjendjen funksionale të gjenomit, ndryshon gjatë gjithë kohës, ndërsa gjenomi mbetet i pandryshuar.
Studimi i proteinave është përdorur prej kohësh për të gjykuar aktivitetin e gjenomave të bimëve. Siç dihet, enzimat e pranishme në të gjitha bimët ndryshojnë në specie dhe varietete individuale në sekuencën e aminoacideve. Enzima të tilla, me të njëjtin funksion, por një sekuencë të ndryshme të aminoacideve individuale, quhen izoenzima. Ato kanë veti të ndryshme fiziko-kimike dhe imunologjike (pesha molekulare, ngarkesa), të cilat mund të zbulohen duke përdorur kromatografi ose elektroforezë. Për shumë vite, këto metoda janë përdorur me sukses për të studiuar të ashtuquajturin polimorfizëm gjenetik, domethënë ndryshimet midis organizmave, varieteteve, popullatave, specieve, në veçanti grurit dhe formave të ngjashme të drithërave. Megjithatë, kohët e fundit, për shkak të zhvillimit të shpejtë të metodave të analizës së ADN-së, duke përfshirë sekuencën, studimi i polimorfizmit të proteinave është zëvendësuar nga studimi i polimorfizmit të ADN-së. Megjithatë, një metodë e rëndësishme dhe e besueshme për analizën gjenetike, përzgjedhjen dhe prodhimin e farës së bimëve bujqësore, mbetet studimi i drejtpërdrejtë i spektrave të proteinave ruajtëse (prolamina, gliadina, etj.), të cilat përcaktojnë vetitë kryesore ushqyese të drithërave.
Njohja e gjeneve, mekanizmave të shprehjes dhe rregullimit të tyre është jashtëzakonisht e rëndësishme për zhvillimin e bioteknologjisë dhe prodhimin e bimëve transgjenike. Dihet se sukseset mbresëlënëse në këtë fushë shkaktojnë një reagim të paqartë nga komuniteti mjedisor dhe mjekësor. Sidoqoftë, ekziston një fushë e bioteknologjisë së bimëve ku këto frikë, nëse jo plotësisht të pabaza, atëherë, në çdo rast, duket se kanë pak rëndësi. Po flasim për krijimin e impianteve industriale transgjenike që nuk përdoren si produkte ushqimore. India së fundmi korri prodhimin e parë të pambukut transgjenik që është rezistent ndaj një sërë sëmundjesh. Ka informacione për futjen e gjeneve speciale që kodojnë proteinat e pigmentit në gjenomën e pambukut dhe prodhimin e fibrave të pambukut që nuk kërkojnë ngjyrosje artificiale. Një tjetër kulturë industriale që mund të jetë objekt i inxhinierisë gjenetike efektive është liri. Përdorimi i tij si një alternativë ndaj pambukut për lëndët e para të tekstilit është diskutuar kohët e fundit. Ky problem është jashtëzakonisht i rëndësishëm për vendin tonë, i cili ka humbur burimet e veta të pambukut të papërpunuar.
PERSPEKTIVAT PËR STUDIMIN E GJENOMËVE TË BIMËVE
Natyrisht, studimet strukturore të gjenomave të bimëve do të bazohen në qasjet dhe metodat e gjenomikës krahasuese, duke përdorur si material kryesor rezultatet e deshifrimit të gjenomave të Arabidopsis dhe orizit. Një rol të rëndësishëm në zhvillimin e gjenomikës krahasuese të bimëve do të luajë padyshim informacioni që herët a vonë do të sigurohet nga sekuenca totale (e përafërt) e gjenomave të bimëve të tjera. Në këtë rast, gjenomika krahasuese e bimëve do të bazohet në vendosjen e marrëdhënieve gjenetike midis lokacioneve individuale dhe kromozomeve që i përkasin gjenomave të ndryshme. Ne do të fokusohemi jo aq në gjenomikën e përgjithshme të bimëve, sa në gjenomikën selektive të lokacioneve individuale kromozomale. Për shembull, kohët e fundit është treguar se gjeni përgjegjës për vernalizimin ndodhet në vendndodhjen VRn-AI të kromozomit hexaploid të grurit 5A dhe vendndodhjen Hd-6 të kromozomit 3 të orizit.
Zhvillimi i këtyre studimeve do të jetë një shtysë e fuqishme për identifikimin, izolimin dhe renditjen e shumë gjeneve të bimëve funksionalisht të rëndësishme, në veçanti, gjeneve përgjegjëse për rezistencën ndaj sëmundjeve, rezistencën ndaj thatësirës dhe përshtatshmërinë ndaj kushteve të ndryshme të rritjes. Gjithnjë e më shumë do të përdoret gjenomika funksionale, bazuar në zbulimin masiv (screening) të gjeneve që funksionojnë në bimë.
Mund të parashikojmë përmirësime të mëtejshme të teknologjive kromozomale, kryesisht metodën e mikrodiseksionit. Përdorimi i tij zgjeron në mënyrë dramatike mundësitë e kërkimit gjenomik pa kërkuar kosto të mëdha, siç është, për shembull, sekuenca totale e gjenomit. Metoda e lokalizimit në kromozomet e bimëve të gjeneve individuale me ndihmën e hibridizimit do të përhapet më tej. në vend. Për momentin, aplikimi i tij është i kufizuar nga numri i madh i sekuencave të përsëritura në gjenomën e bimëve, dhe ndoshta nga veçoritë e organizimit strukturor të kromozomeve bimore.
Teknologjitë kromozomale do të kenë një rëndësi të madhe për gjenominë evolucionare të bimëve në të ardhmen e parashikueshme. Këto teknologji relativisht të lira bëjnë të mundur vlerësimin e shpejtë të ndryshueshmërisë brenda dhe ndërspecifike, studimin e gjenomave komplekse alopoliploide të grurit tetraploid dhe hexaploid, triticale; të analizojë proceset evolucionare në nivel kromozomik; të hetojë formimin e gjenomave sintetike dhe futjen (introgresionin) e materialit gjenetik të huaj; identifikojnë marrëdhëniet gjenetike midis kromozomeve individuale të llojeve të ndryshme.
Studimi i kariotipit të bimës duke përdorur metoda klasike citogjenetike, të pasuruara me analiza biologjike molekulare dhe teknologji kompjuterike, do të përdoret për të karakterizuar gjenomin. Kjo është veçanërisht e rëndësishme për studimin e stabilitetit dhe ndryshueshmërisë së kariotipit në nivel jo vetëm të organizmave individualë, por edhe të popullatave, varieteteve dhe specieve. Së fundi, është e vështirë të imagjinohet se si mund të vlerësohet numri dhe spektri i rirregullimeve kromozomale (aberracionet, urat) pa përdorimin e metodave të ngjyrosjes diferenciale. Studime të tilla janë jashtëzakonisht premtuese për monitorimin e mjedisit sipas gjendjes së gjenomit të bimëve.
Në Rusinë moderne, sekuenca e drejtpërdrejtë e gjenomave të bimëve nuk ka gjasa të kryhet. Një punë e tillë, që kërkon investime të mëdha, është përtej fuqisë së ekonomisë sonë aktuale. Ndërkohë, të dhënat për strukturën e gjenomave të Arabidopsis dhe orizit, të marra nga shkenca botërore dhe të disponueshme në bankat ndërkombëtare të të dhënave, janë të mjaftueshme për zhvillimin e gjenomikës së bimëve vendase. Mund të parashikohet zgjerimi i studimeve të gjenomave të bimëve bazuar në qasjet e gjenomikës krahasuese për zgjidhjen e problemeve specifike të mbarështimit dhe prodhimit të bimëve, si dhe për të studiuar origjinën e llojeve të ndryshme bimore me rëndësi të madhe ekonomike.
Mund të supozohet se qasjet gjenomike si tipizimi gjenetik (RELF, RAPD, analizat AFLP, etj.), të cilat janë mjaft të përballueshme për buxhetin tonë, do të përdoren gjerësisht në praktikën e mbarështimit vendas dhe në prodhimin e bimëve. Paralelisht me metodat e drejtpërdrejta për përcaktimin e polimorfizmit të ADN-së, qasjet e bazuara në studimin e polimorfizmit të proteinave, kryesisht proteinat ruajtëse të drithërave, do të përdoren për të zgjidhur problemet e gjenetikës dhe mbarështimit të bimëve. Teknologjitë kromozomale do të përdoren gjerësisht. Ato janë relativisht të lira, zhvillimi i tyre kërkon investime mjaft të moderuara. Në fushën e studimeve të kromozomeve, shkenca vendase nuk është inferiore ndaj botës.
Duhet theksuar se shkenca jonë ka dhënë një kontribut të rëndësishëm në formimin dhe zhvillimin e gjenomikës së bimëve [ , ].
Rolin themelor e luajti N.I. Vavilov (1887-1943).
Në biologjinë molekulare dhe gjenomikën e bimëve, kontributi pionier i A.N. Belozersky (1905-1972).
Në fushën e studimeve kromozomale, është e nevojshme të theksohet puna e gjenetistit të shquar S.G. Navashin (1857-1930), i cili së pari zbuloi kromozomet satelitore në bimë dhe vërtetoi se është e mundur të bëhet dallimi midis kromozomeve individuale sipas veçorive të morfologjisë së tyre.
Një tjetër klasik i shkencës ruse G.A. Levitsky (1878-1942) përshkroi në detaje kromozomet e thekrës, grurit, elbit, bizeles dhe panxharit të sheqerit, futi termin "kariotip" në shkencë dhe zhvilloi doktrinën e tij.
Specialistët modernë, duke u mbështetur në arritjet e shkencës botërore, mund të japin një kontribut të rëndësishëm në zhvillimin e mëtejshëm të gjenetikës dhe gjenomisë së bimëve.
Autori shpreh falënderimet e tij të përzemërta për Akademik Yu.P. Altukhov për diskutimin kritik të artikullit dhe këshilla të vlefshme.Puna e ekipit të kryesuar nga autori i artikullit u mbështet nga Fondacioni Rus për Kërkime Bazë (grantet nr. 99-04-48832; 00-04-49036; 00-04-81086), Programi i Presidentit të Federata Ruse për të mbështetur shkollat shkencore (grantet nr. 00-115 -97833 dhe NSh-1794.2003.4) dhe Programin e Akademisë Ruse të Shkencave "Shenuesit molekularë gjenetikë dhe kromozomalë në zhvillimin e metodave moderne të mbarështimit dhe prodhimit të farës" .
LITERATURA
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Hyrje në gjenomikën e bimëve // Biologjia molekulare. 2001. V. 35. S. 339-348. 2. Stilolaps E. Bonanza për gjenomikën e bimëve // Shkencë. 1998. V. 282. F. 652-654. 3. Gjenomika e bimëve, Proc. Natl. Akad. shkenca. SHBA. 1998. V. 95. P. 1962-2032. 4. Karteli N.A. dhe etj. Gjenetika. Fjalor Enciklopedik. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferencimi i gjenomit në Aegilops. 1. Shpërndarja e sekuencave të ADN-së shumë të përsëritura në kromozomet e specieve diploide // Gjenomi. 1996. V. 39. F. 293-306. Historia e analizës së kromozomeve // Biol. membranat. 2001. T. 18. S. 164-172.
Gjenet kërcyese
Në mesin e shekullit të kaluar, studiuesja amerikane Barbara McClintock zbuloi gjene të mahnitshme në misër që mund të ndryshojnë në mënyrë të pavarur pozicionin e tyre në kromozome. Tani ato quhen "gjene kërcyese" ose elementë të transpozueshëm (të lëvizshëm). Zbulimi nuk u njoh për një kohë të gjatë, duke i konsideruar elementët e lëvizshëm si një fenomen unik, karakteristik vetëm për misrin. Megjithatë, ishte për këtë zbulim në 1983 që McClintock u shpërblye Çmimi Nobël Sot, gjenet kërcyese janë gjetur në pothuajse të gjitha llojet e kafshëve dhe bimëve të studiuara.
Nga kanë ardhur gjenet hopping, çfarë bëjnë në qelizë, a ka ndonjë përfitim prej tyre? Pse, me prindër të shëndetshëm gjenetikisht, familja e mizës së frutave Drosophila, për shkak të gjeneve kërcyese, mund të prodhojë pasardhës mutantë me një frekuencë të lartë apo edhe të jetë plotësisht pa fëmijë? Cili është roli i gjeneve kërcyese në evolucion?
Duhet thënë se gjenet që sigurojnë funksionimin e qelizave janë të vendosura në kromozome në një renditje të caktuar. Falë kësaj, për shumë lloje të organizmave njëqelizorë dhe shumëqelizorë, u bë e mundur të ndërtoheshin të ashtuquajturat. hartat gjenetike. Megjithatë, ekziston një rend i madhësisë më shumë material gjenetik midis gjeneve sesa në vetvete! Se çfarë roli luan kjo pjesë "çakëll" e ADN-së nuk është vërtetuar plotësisht, por pikërisht këtu gjenden më shpesh elementë të lëvizshëm, të cilët jo vetëm lëvizin vetë, por mund të marrin edhe fragmente fqinje të ADN-së me vete.

Nga vijnë gjenet jumper? Besohet se të paktën disa prej tyre kanë origjinën nga viruset, pasi disa elementë të lëvizshëm janë në gjendje të formojnë grimca virale (për shembull, elementi i lëvizshëm cigan në mizën e frutave Drosophila melanogaster). Disa elemente të transpozueshme shfaqen në gjenom nga të ashtuquajturat transferimi horizontal nga llojet e tjera. Për shembull, konstatohet se celulari endacak-element (i përkthyer në rusisht, ai quhet tramp) Drosophila melanogaster futur në mënyrë të përsëritur në gjenomin e kësaj specie. Ekziston një version që disa rajone rregullatore të ADN-së mund të kenë gjithashtu autonomi dhe një tendencë për "endacak".
çakëll i dobishëm
Nga ana tjetër, shumica e gjeneve kërcyese, pavarësisht emrit, sillen në heshtje, megjithëse përbëjnë një të pestën e materialit gjenetik total. Drosophila melanogaster ose pothuajse gjysma e gjenomit njerëzor.
Teprica e ADN-së, e cila u përmend më lart, ka plusin e vet: ADN-ja e çakëllit (përfshirë elementët e lëvizshëm pasivë) merr goditjen nëse ADN-ja e huaj futet në gjenom. Mundësia që një element i ri të futet në një gjen të dobishëm dhe në këtë mënyrë të prishë funksionimin e tij zvogëlohet nëse ka ADN shumë më të madhe se sa domethënëse.
Një sasi e tepërt e ADN-së është e dobishme në të njëjtën mënyrë si "teprica" e shkronjave në fjalë: ne shkruajmë "Maria Ivanovna" dhe themi "Marivana". Disa nga shkronjat humbasin në mënyrë të pashmangshme, por kuptimi mbetet. I njëjti parim funksionon edhe në nivelin e rëndësisë së aminoacideve individuale në një molekulë proteine-enzimë: vetëm sekuenca e aminoacideve që formon qendrën aktive është rreptësisht konservatore. Kështu, në nivele të ndryshme, teprica rezulton të jetë një lloj buferi që ofron një marzh sigurie për sistemin. Kështu elementët e lëvizshëm që kanë humbur lëvizshmërinë e tyre nuk janë të padobishëm për gjenomin. Siç thonë ata, "nga një dele e hollë edhe një tufë leshi", megjithëse, mbase, një proverb tjetër do të ishte më i përshtatshëm këtu - "çdo bast në një rresht".

Elementët e lëvizshëm që kanë ruajtur aftësinë për të kërcyer lëvizin përgjatë kromozomeve Drosophila me një frekuencë prej 10-2-10-5 për gjen për gjeneratë, në varësi të llojit të elementit, sfondit gjenetik dhe kushtet e jashtme. Kjo do të thotë se një nga njëqind gjenet kërcyese në një qelizë mund të ndryshojë pozicionin e tij pas ndarjes së ardhshme të qelizave. Si rezultat, pas disa brezash, shpërndarja e elementeve të transpozueshme përgjatë kromozomit mund të ndryshojë shumë ndjeshëm.
Është i përshtatshëm për të studiuar një shpërndarje të tillë në kromozomet politen (multifilamentoze) nga gjëndrat e pështymës së larvave Drosophila. Këto kromozome janë shumë herë më të trasha se ato normale, duke e bërë shumë më të lehtë ekzaminimin e tyre nën një mikroskop. Si krijohen këto kromozome? Në qelizat e gjëndrave të pështymës, ADN-ja e secilit prej kromozomeve shumëfishohet, si në ndarjen normale të qelizave, por vetë qeliza nuk ndahet. Si rezultat, numri i qelizave në gjëndër nuk ndryshon, por në 10-11 cikle, disa mijëra fije identike të ADN-së grumbullohen në secilin kromozom.

Pjesërisht për shkak të kromozomeve të politenit, gjenet kërcyese në Drosophila kuptohen më mirë se në metazoanët e tjerë. Si rezultat i këtyre studimeve, rezultoi se edhe brenda të njëjtës popullatë Drosophila është e vështirë të gjenden dy individë që kanë kromozome me të njëjtën shpërndarje të elementeve të lëvizshëm. Nuk është rastësi që shumica e mutacioneve spontane në Drosophila besohet se shkaktohen nga lëvizja e këtyre "hoppers".
Pasojat mund të ndryshojnë...
Bazuar në efektin e tyre në gjenom, elementët aktivë të transpozueshëm mund të ndahen në disa grupe. Disa prej tyre kryejnë funksione jashtëzakonisht të rëndësishme dhe të dobishme për gjenomin. Për shembull, telomerike ADN-ja, e vendosur në skajet e kromozomeve, në Drosophila përbëhet vetëm nga elementë të veçantë të lëvizshëm. Kjo ADN është jashtëzakonisht e rëndësishme - humbja e saj sjell humbjen e të gjithë kromozomit në procesin e ndarjes së qelizave, gjë që çon në vdekjen e qelizave.
Elementë të tjerë të lëvizshëm janë "dëmtues" të plotë. Të paktën si të tilla konsiderohen ky moment. Për shembull, elementët e transpozueshëm të klasës R2 mund të futen posaçërisht në gjenet e artropodëve që kodojnë një nga proteinat e ribozomeve - "fabrikat" qelizore për sintezën e proteinave. Individët me çrregullime të tilla mbijetojnë vetëm sepse vetëm një pjesë e gjeneve të shumta që kodojnë këto proteina dëmtohet në gjenom.

Ka edhe elementë të tillë të lëvizshëm që lëvizin vetëm në indet riprodhuese që prodhojnë qeliza germinale. Kjo shpjegohet me faktin se në inde të ndryshme i njëjti element i lëvizshëm mund të prodhojë gjatësi dhe funksione të ndryshme të molekulës protein-enzimë të nevojshme për lëvizjen.
Një shembull i këtij të fundit është elementi P Drosophila melanogaster, e cila u fut në popullatat e saj natyrore me transferim horizontal nga një specie tjetër e Drosophila jo më shumë se njëqind vjet më parë. Megjithatë, nuk ka pothuajse një popullsi në Tokë tani. Drosophila melanogaster, në të cilin nuk do të kishte asnjë element P. Në të njëjtën kohë, duhet të theksohet se shumica e kopjeve të tij janë me defekt, për më tepër, i njëjti version i defektit u gjet pothuajse kudo. Roli i këtij të fundit në gjenom është i veçantë: është "jotolerant" ndaj shokëve të tij dhe luan rolin e një shtypësi, duke bllokuar lëvizjen e tyre. Pra, mbrojtja e gjenomit të Drosophila nga kërcimet e "alienit" mund të kryhet pjesërisht nga derivatet e tij.
Gjëja kryesore është të zgjidhni prindërit e duhur!
Shumica e kërcimeve të elementëve celularë nuk ndikojnë pamjen Drosophila, sepse bie në ADN-në e çakëllit, por ka situata të tjera kur aktiviteti i tyre rritet në mënyrë dramatike.

Mjaft e çuditshme, faktori më i fuqishëm që nxit lëvizjen e gjeneve kërcyese është prindërimi i dobët. Për shembull, çfarë ndodh nëse kryqëzoni femra nga një popullatë laboratorike Drosophila melanogaster, të cilat nuk kanë një element P (sepse paraardhësit e tyre u kapën nga natyra rreth njëqind vjet më parë), me meshkujt që mbanin një element P? Në hibride, për shkak të lëvizjes së shpejtë të elementit të lëvizshëm, mund të shfaqen një numër i madh i çrregullimeve të ndryshme gjenetike. Ky fenomen, i quajtur disgjenezë hibride, shkaktohet nga mungesa e një represori në citoplazmën e nënës që ndalon lëvizjen e elementit të lëvizshëm.
Kështu, nëse dhëndërit nga popullsia A dhe nuset nga popullsia B mund të krijojnë familje të mëdha, atëherë nuk është gjithmonë e kundërta. Një familje me prindër të shëndetshëm gjenetikisht mund të prodhojë një numër të madh pasardhëssh mutant ose jopjellor, apo edhe të jetë pa fëmijë nëse babai dhe nëna kanë një grup të ndryshëm elementësh të lëvizshëm në gjenom. Veçanërisht shumë shkelje shfaqen nëse eksperimenti kryhet në një temperaturë prej 29 ° C. Ndikimi i faktorëve të jashtëm, të mbivendosur në sfondin gjenetik, rrit efektin e mospërputhjes së gjenomit, megjithëse vetëm këta faktorë (madje edhe rrezatimi jonizues) nuk janë të aftë. duke shkaktuar një lëvizje kaq masive të elementëve të lëvizshëm.
Ngjarje të ngjashme në Drosophila melanogaster mund të ndodhë me pjesëmarrjen e familjeve të tjera të elementëve të lëvizshëm.
Evolucioni "Mobile".
Gjenomi qelizor mund të shihet si një lloj ekosistemi i anëtarëve të përhershëm dhe të përkohshëm, ku fqinjët jo vetëm që bashkëjetojnë, por edhe ndërveprojnë me njëri-tjetrin. Ndërveprimi i gjeneve të bujtësit me elementët e transpozueshëm ende nuk është kuptuar mirë, por mund të citohen shumë rezultate - nga vdekja e një organizmi në rast të dëmtimit të një gjeni të rëndësishëm deri në rivendosjen e funksioneve të dëmtuara më parë.
Ndodh që vetë gjenet kërcyese ndërveprojnë me njëri-tjetrin. Kështu, njihet një fenomen që i ngjan imunitetit, kur një element i lëvizshëm nuk mund të futet në afërsi të një elementi ekzistues. Sidoqoftë, jo të gjithë elementët celularë janë kaq delikatë: për shembull, elementët P mund të futen lehtësisht në njëri-tjetrin dhe t'i nxjerrin vëllezërit e tyre nga loja.

Përveç kësaj, ekziston një lloj vetë-rregullimi i numrit të elementeve të transpozueshme në gjenom. Fakti është se elementët e lëvizshëm mund të shkëmbejnë rajone homologe me njëri-tjetrin - ky proces quhet rikombinim. Si rezultat i një ndërveprimi të tillë, elementët celularë, në varësi të orientimit të tyre, mund të humbasin ( fshirje) ose zgjerohet ( përmbysja) fragmente të ADN-së së bujtësit të vendosura ndërmjet tyre. Nëse një pjesë e rëndësishme e kromozomit humbet, gjenomi do të vdesë. Në rastin e një përmbysjeje ose një delecioni të vogël, krijohet diversiteti i kromozomeve, i cili konsiderohet një kusht i domosdoshëm për evolucionin.
Nëse ndodhin rikombinime midis elementeve të lëvizshëm të vendosur në kromozome të ndryshme, atëherë si rezultat formohen rirregullime kromozomike, të cilat, gjatë ndarjeve të mëvonshme të qelizave, mund të çojnë në një çekuilibër në gjenom. Dhe një gjenom i pabalancuar, si një buxhet i pabalancuar, është shumë i ndarë keq. Pra, vdekja e gjenomave të pasuksesshme është një nga arsyet pse elementët aktivë të transpozueshëm nuk i vërshojnë kromozomet pa kufi.
Lind një pyetje e natyrshme: sa i rëndësishëm është kontributi i elementëve të lëvizshëm në evolucion? Së pari, shumica e elementeve të transpozueshme futen, përafërsisht, aty ku duhet, si rezultat i të cilave mund të dëmtojnë ose ndryshojnë strukturën ose rregullimin e gjenit në të cilin futen. Pastaj seleksionimi natyror fshin mënjanë opsionet e pasuksesshme dhe opsionet e suksesshme me veti adaptive fiksohen.

Nëse pasojat e futjes së një elementi të transpozueshëm rezultojnë të jenë neutrale, atëherë ky variant mund të ruhet në popullatë, duke siguruar një lloj diversiteti në strukturën e gjenit. Kjo mund të jetë e dobishme kur kushte të pafavorshme. Teorikisht, me një lëvizje masive të elementeve të lëvizshëm, mutacionet mund të shfaqen në shumë gjene në të njëjtën kohë, gjë që mund të jetë shumë e dobishme në një ndryshim të mprehtë të kushteve të ekzistencës.
Pra, për të përmbledhur: ka shumë elementë të lëvizshëm në gjenom dhe ata janë të ndryshëm; ato mund të ndërveprojnë si me njëri-tjetrin ashtu edhe me gjenet strehuese; mund të jetë i dëmshëm dhe i pazëvendësueshëm. Paqëndrueshmëria e gjenomit e shkaktuar nga lëvizja e elementeve të lëvizshëm mund të përfundojë në tragjedi për një individ, por aftësia për të ndryshuar shpejt është një kusht i domosdoshëm për mbijetesën e një popullate ose specie. Kjo krijon diversitet, i cili është baza për përzgjedhjen natyrore dhe transformimet e mëvonshme evolucionare.
Ju mund të bëni një farë analogjie midis gjeneve kërcyese dhe emigrantëve: disa emigrantë ose pasardhës të tyre bëhen qytetarë të barabartë, të tjerëve u jepet leje qëndrimi dhe të tjerëve - ata që nuk respektojnë ligjet - deportohen ose burgosen. Dhe migrimet masive të popujve mund të ndryshojnë shpejt vetë shtetin.
Letërsia
Ratner V. A., Vasilyeva L. A. Induksioni i transpozimeve të elementeve gjenetike të lëvizshme nga ndikimet stresuese. Lidhja ruse. 2000.
Gvozdev V. A. ADN eukariote lëvizëse // Revista Arsimore Soros. 1998. Nr. 8.





