Dešifrovanje ljudskog genoma. Medicinska genetika Prilikom dešifriranja genoma Drosophila, ustanovljeno je da
Povodom 50. godišnjice otkrića strukture DNK
A.V. Zelenin
BILJNI GENOM
A. V. Zelenin
Zelenin Aleksandar Vladimirovič- d.b.n.,
šef laboratorije Instituta za molekularnu biologiju. V.A. Engelhardt RAS.
Impresivna dostignuća programa "Ljudski genom", kao i uspeh rada na dešifrovanju takozvanih ekstra malih (virusi), malih (bakterije, kvasac) i srednjih (okrugli crvi, Drosophila) genoma, omogućili su premještanje na veliku studiju velikih i ekstra velikih biljnih genoma. Hitna potreba za detaljnim proučavanjem genoma ekonomski najvažnijih biljaka naglašena je na skupu o biljnoj genomici održanom 1997. godine u Sjedinjenim Državama [ , ]. Tokom godina koje su prošle od tog vremena, postignuti su nesumnjivi uspjesi u ovoj oblasti. 2000. godine pojavila se publikacija o potpunom sekvenciranju (određivanje linearne sekvence nukleotida cjelokupne nuklearne DNK) genoma male gorušice - Arabidopsis, 2001. godine - o preliminarnom (nacrtu) sekvenciranju genoma riže. Radovi na sekvenciranju genoma velikih i super velikih biljaka (kukuruz, raž, pšenica) su više puta izvještavani, međutim, ti izvještaji nisu sadržavali konkretne informacije i više su bili po prirodi deklaracija namjere.
Pretpostavlja se da će dekodiranje biljnih genoma otvoriti široke perspektive za nauku i praksu. Prije svega, identifikacija novih gena i lanac njihove genetske regulacije značajno će povećati produktivnost biljaka korištenjem biotehnoloških pristupa. Otkrivanjem, izolacijom, reprodukcijom (kloniranjem) i sekvenciranjem gena odgovornih za tako važne funkcije biljnog organizma kao što su reprodukcija i produktivnost, procesi varijabilnosti, otpornost na štetne faktore okoline, kao i homologno uparivanje hromozoma, dolazi do pojave povezane su nove mogućnosti za unapređenje procesa uzgoja. Konačno, izolovani i klonirani geni se mogu koristiti za dobijanje transgenih biljaka sa fundamentalno novim svojstvima i za analizu mehanizama regulacije aktivnosti gena.
Važnost proučavanja biljnih genoma naglašava i činjenica da je do sada broj lokaliziranih, kloniranih i sekvenciranih biljnih gena mali i varira, prema različitim procjenama, između 800 i 1200. To je 10-15 puta manje nego za na primjer, kod ljudi.
Sjedinjene Američke Države ostaju nesumnjivi lider u istraživanju genoma biljaka velikih razmjera, iako se intenzivne studije genoma riže provode u Japanu, te u poslednjih godina iu Kini. U dešifrovanju genoma Arabidopsis, pored američkih laboratorija, aktivno su učestvovale i evropske istraživačke grupe. Očigledno vodstvo Sjedinjenih Država izaziva ozbiljnu zabrinutost evropskih naučnika, što su jasno izrazili na skupu pod značajnim naslovom "Izgledi za genomiku u postgenomskoj eri", održanom krajem 2000. godine u Francuskoj. Napredak američke nauke u proučavanju genoma poljoprivrednih biljaka i stvaranju transgenih biljnih formi, prema evropskim naučnicima, prijeti da će u ne tako dalekoj budućnosti (dvije do pet decenija), kada će porast populacije staviti čovječanstvo u lice opšteg prehrambena kriza, evropska ekonomija i nauka će postati zavisni od američke tehnologije. S tim u vezi, najavljeno je stvaranje francusko-njemačkog naučnog programa za proučavanje biljnih genoma ("Plantgene") iu njega su uložena značajna ulaganja.
Očigledno je da bi problemi biljne genomike trebali privući veliku pažnju ruskih naučnika i organizatora nauke, ali i vlasti, jer se ne radi samo o naučnom prestižu, već i o nacionalnoj sigurnosti zemlje. Za deceniju ili dve, hrana će postati najvažniji strateški resurs.
TEŠKOĆE U PROUČAVANJU BILJNIH GENOMA
Proučavanje genoma biljaka mnogo je teži zadatak od proučavanja genoma ljudi i drugih životinja. To je zbog sljedećih okolnosti:
ogromne veličine genoma, posežući za određene vrste biljke od desetina, pa čak i stotina milijardi baznih parova (bp): genomi glavnih ekonomski važnih biljaka (osim riže, lana i pamuka) su ili bliski ljudskom genomu ili višestruko veći (tabela);HROMOSOMSKA STUDIJA GENOMAOštre fluktuacije u broju hromozoma u različitim biljkama - od dva kod nekih vrsta do nekoliko stotina u drugih, i nije moguće utvrditi strogu korelaciju između veličine genoma i broja hromozoma;
Obilje poliploida (koji sadrže više od dva genoma po ćeliji) formira se sa sličnim, ali ne identičnim genomima (alpoliploidija);
Ekstremno obogaćivanje biljnih genoma (do 99%) "beznačajne" (nekodirajuće, odnosno ne sadrži gene) DNK, što otežava spajanje sekvencioniranih fragmenata (poređanje u ispravnom redoslijedu) u zajednički DNK region velike veličine (kontig);
Nepotpuno (u poređenju sa genomima drozofile, čoveka i miša) morfološko, genetsko i fizičko mapiranje hromozoma;
Praktična nemogućnost izolacije pojedinačnih hromozoma u čistom obliku korišćenjem metoda koje se obično koriste u tu svrhu za ljudske i životinjske hromozome (razvrstavanje u struji i upotreba ćelijskih hibrida);
Poteškoća hromozomskog mapiranja (određivanje lokacije na hromozomu) pojedinačnih gena pomoću hibridizacije in situ, kako zbog visokog sadržaja "beznačajne" DNK u biljnim genomima, tako i zbog posebnosti strukturne organizacije biljnih hromozoma;
Evolucijska udaljenost biljaka od životinja, što ozbiljno otežava korištenje informacija dobivenih sekvenciranjem genoma ljudi i drugih životinja za proučavanje genoma biljaka;
Dug proces reprodukcije većine biljaka, što značajno usporava njihovu genetsku analizu.
Kromosomske (citogenetske) studije genoma općenito, a posebno biljaka duga istorija. Termin "genom" predložen je za označavanje haploidnog (jednostrukog) skupa hromozoma sa genima koji se nalaze u njima u prvoj četvrtini 20. veka, odnosno mnogo pre uspostavljanja uloge DNK kao nosioca genetske informacije. .
Opis genoma novog, ranije genetski neistraženog višećelijskog organizma obično počinje proučavanjem i opisom kompletnog skupa njegovih hromozoma (kariotipa). To se, naravno, odnosi i na biljke, od kojih veliki broj nije ni počeo da se proučava.
Već na početku istraživanja hromozoma upoređivani su genomi srodnih biljnih vrsta na osnovu analize mejotičke konjugacije (kombinacije homolognih hromozoma) u interspecifičnim hibridima. Tokom proteklih 100 godina, mogućnosti analize hromozoma su se dramatično proširile. Sada se za karakterizaciju biljnih genoma koriste naprednije tehnologije: razne varijante takozvanog diferencijalnog bojenja, koje omogućava identifikaciju pojedinačnih hromozoma prema morfološkim karakteristikama; hibridizacija in situ omogućavajući lokalizaciju specifičnih gena na hromozomima; biohemijsko istraživanjećelijskih proteina (elektroforeza i imunohemija) i, konačno, skup metoda zasnovanih na analizi hromozomske DNK do njenog sekvenciranja.

Rice. jedan. Kariotipovi žitarica a - raž (14 hromozoma), b - durum pšenica (28 hromozoma), c - meka pšenica (42 hromozoma), d - ječam (14 hromozoma)Dugi niz godina proučavaju se kariotipovi žitarica, prvenstveno pšenice i raži. Zanimljivo je to različite vrste kod ovih biljaka, broj hromozoma je različit, ali je uvijek višestruk od sedam. Pojedine vrste žitarica mogu se pouzdano prepoznati po kariotipu. Na primjer, genom raži sastoji se od sedam parova velikih hromozoma sa intenzivno obojenim heterokromatskim blokovima na njihovim krajevima, koji se često nazivaju segmenti ili trake (slika 1a). Genomi pšenice već imaju 14 i 21 par hromozoma (sl. 1, b, c), a distribucija heterokromatskih blokova u njima nije ista kao u hromozomima raži. Pojedinačni genomi pšenice, označeni A, B i D, također se međusobno razlikuju. Povećanje broja hromozoma sa 14 na 21 dovodi do nagle promjene svojstava pšenice, što se ogleda u njihovim nazivima: durum, ili testenina, pšenica i mekana, ili hleb, pšenica. D gen, koji sadrži gene za proteine glutena, koji daje testu takozvanu klijavost, odgovoran je za sticanje visokih pekarskih svojstava mekom pšenicom. Upravo ovom genomu se pridaje posebna pažnja u selekcijskom poboljšanju hljebne pšenice. Druga žitarica sa 14 hromozoma, ječam (slika 1, d), obično se ne koristi za pravljenje hleba, ali je glavna sirovina za proizvodnju uobičajenih proizvoda kao što su pivo i viski.
Intenzivno se istražuju hromozomi nekih samoniklih biljaka koji se koriste za poboljšanje kvaliteta najvažnijih poljoprivrednih vrsta, kao što je divlji srodnik pšenice - Aegilops. Nove biljne forme nastaju ukrštanjem (slika 2) i selekcijom. Posljednjih godina značajno poboljšanje u istraživačkim metodama omogućilo je početak proučavanja biljnih genoma, čije su karakteristike kariotipova (uglavnom mala veličina kromosoma) činile ranije nedostupnim za kromosomsku analizu. Dakle, tek nedavno su prvi put identificirani svi hromozomi pamuka, kamilice i lana.

Rice. 2. Kariotipovi pšenice i hibrid pšenice sa Aegilopsom
a - heksaploidna meka pšenica ( Triticum astivum), koji se sastoji od A, B i O genoma; b - tetraploidna pšenica ( Triticum timopheevi), koji se sastoji od A i G genoma. sadrži gene za otpornost na većinu bolesti pšenice; c - hibridi Triticum astivum X Triticum timopheevi otporan na pepelnicu i rđu, jasno je vidljiva zamjena dijela hromozomaPRIMARNA STRUKTURA DNK
Razvojem molekularne genetike proširio se i sam koncept genoma. Sada se ovaj pojam tumači i u klasičnom hromozomskom i u modernom molekularnom smislu: cjelokupni genetski materijal pojedinačnog virusa, ćelije i organizma. Naravno, nakon proučavanja kompletne primarne strukture genoma (kako se često naziva kompletan linearni niz baza nukleinskih kiselina) brojnih mikroorganizama i ljudi, postavilo se pitanje sekvenciranja biljnog genoma.
Od brojnih biljnih organizama, dva su odabrana za proučavanje - Arabidopsis, koji predstavlja klasu dvosupnica (veličina genoma 125 miliona bp), i pirinač iz klase jednosupnica (420-470 miliona bp). Ovi genomi su mali u poređenju sa drugim biljnim genomima i sadrže relativno malo segmenata DNK koji se ponavljaju. Takve karakteristike su davale nadu da će odabrani genomi biti dostupni za relativno brzo određivanje njihove primarne strukture.

Rice. 3. Arabidopsis - mala gorušica - mala biljka iz porodice krstaša ( Brassicaceae). Na prostoru jednakoj površini jednoj stranici našeg časopisa, možete uzgajati do hiljadu pojedinačnih organizama Arabidopsis.Razlog odabira Arabidopsisa nije bila samo mala veličina njegovog genoma, već i mala veličina organizma, što ga čini lakim za uzgoj u laboratoriji (slika 3). Uzeli smo u obzir njegov kratak reproduktivni ciklus, koji omogućava brzo provođenje eksperimenata ukrštanja i selekcije, detaljno proučavanu genetiku, lakoću manipulacije s promjenjivim uvjetima uzgoja (promjena sastava soli u tlu, dodavanje različitih hranjivih tvari, itd.) i ispitivanje djelovanja na biljke raznih mutagenih faktora i patogena (virusa, bakterija, gljivica). Arabidopsis nema ekonomsku vrijednost, stoga je njen genom, zajedno sa genomom miša, nazvan referentnim, ili, manje tačno, modelom.*
* Pojava pojma "model genoma" u ruskoj literaturi rezultat je netačnog prijevoda engleske fraze model genoma. Riječ "model" znači ne samo pridjev "model", već i imenicu "uzorak", "standard", "model". Ispravnije bi bilo govoriti o uzorku genoma ili referentnom genomu.Intenzivan rad na sekvenciranju genoma Arabidopsis započeo je 1996. godine od strane međunarodnog konzorcijuma koji je uključivao naučne institucije i istraživačke grupe iz SAD-a, Japana, Belgije, Italije, Velike Britanije i Njemačke. U decembru 2000. godine postale su dostupne opsežne informacije koje sumiraju određivanje primarne strukture genoma Arabidopsis. Za sekvenciranje je korištena klasična ili hijerarhijska tehnologija: prvo su proučavani pojedinačni mali dijelovi genoma od kojih su sastavljeni veći dijelovi (kontigi), a u završnoj fazi struktura pojedinačnih hromozoma. Nuklearna DNK genoma Arabidopsis raspoređena je na pet hromozoma. Godine 1999. objavljeni su rezultati sekvenciranja dva hromozoma, a pojavljivanje u štampi informacija o primarnoj strukturi preostala tri dovršilo je sekvenciranje cijelog genoma.
Od 125 miliona baznih parova utvrđena je primarna struktura od 119 miliona, što je 92% ukupnog genoma. Ispostavilo se da je samo 8% genoma Arabidopsis koji sadrži velike blokove repetitivnih segmenata DNK nepristupačno za proučavanje. U pogledu kompletnosti i temeljitosti sekvenciranja eukariotskog genoma, Arabidopsis ostaje među prva tri šampiona zajedno sa jednoćelijskim kvascem. Saccharomyces cerevisiae i višećelijski organizam Caenorhabditis elegance(vidi tabelu).
Oko 15.000 pojedinačnih gena koji kodiraju proteine pronađeno je u genomu Arabidopsis. Otprilike 12.000 njih sadržano je kao dvije kopije po haploidnom (jednom) genomu, tako da ukupan broj gena je 27 hiljada Broj gena kod Arabidopsisa se ne razlikuje mnogo od broja gena u organizmima kao što su ljudi i miševi, ali je veličina njegovog genoma 25-30 puta manja. Ova okolnost povezana je sa važnim karakteristikama u strukturi pojedinačnih gena Arabidopsis i cjelokupnoj strukturi njenog genoma.
Geni Arabidopsis su kompaktni, sadrže samo nekoliko egzona (protein-kodirajućih regija) razdvojenih kratkim (oko 250 bp) nekodirajućim DNK segmentima (intronima). Intervali između pojedinačnih gena su u prosjeku 4600 parova baza. Poređenja radi, ističemo da ljudski geni sadrže mnogo desetina, pa čak i stotina egzona i introna, a međugenske regije imaju veličine od 10 hiljada parova baza ili više. Pretpostavlja se da je prisustvo malog kompaktnog genoma doprinijelo evolucijskoj stabilnosti Arabidopsis, budući da je njegova DNK u manjoj mjeri postala meta raznih štetnih agenasa, posebno za uvođenje repetitivnih fragmenata DNK (transpozona) sličnih virusu. u genom.
Od ostalih molekularnih karakteristika genoma Arabidopsis, treba napomenuti da su egzoni obogaćeni gvaninom i citozinom (44% u egzonima i 32% u intronima) u poređenju sa životinjskim genima, kao i prisustvo dvostruko ponovljenih (dupliciranih) gena. . Pretpostavlja se da je do takvog udvostručavanja došlo kao rezultat četiri istovremena događaja, koji se sastoje od udvostručavanja (ponavljanja) dijela gena Arabidopsis, odnosno fuzije srodnih genoma. Ovi događaji, koji su se odigrali prije 100-200 miliona godina, manifestacija su općeg trenda poliploidizacije (višestruko povećanje broja genoma u organizmu), koji je karakterističan za biljne genome. Međutim, neke činjenice pokazuju da duplicirani geni u Arabidopsisu nisu identični i funkcioniraju drugačije, što može biti povezano s mutacijama u njihovim regulatornim regijama.
Riža je postala još jedan predmet kompletnog sekvenciranja DNK. Genom ove biljke je također mali (12 hromozoma, što daje ukupno 420-470 miliona bp), samo 3,5 puta veći od genoma Arabidopsis. Međutim, za razliku od Arabidopsisa, riža je od velike ekonomske važnosti, budući da je osnova ishrane više od polovine čovječanstva, dakle, ne samo milijarde potrošača, već i višemilionska armija ljudi aktivno uključenih u veoma naporan proces njenog kultivacije su od vitalnog značaja za poboljšanje svojih svojstava.
Neki istraživači su počeli proučavati genom riže još 1980-ih, ali su te studije dostigle ozbiljne razmjere tek 1990-ih. Godine 1991. u Japanu je kreiran program za dešifrovanje strukture genoma riže, objedinjujući napore mnogih istraživačkih grupa. Na osnovu ovog programa 1997. godine organizovan je Međunarodni projekat genoma riže. Njegovi učesnici odlučili su da koncentrišu svoje napore na sekvencioniranje jedne od podvrsta riže ( Oriza sativajaponica), u čijem proučavanju je do tada već postignut značajan napredak. Ozbiljan podsticaj i, slikovito rečeno, zvijezda vodilja za takav rad bio je program "Ljudski genom".
U okviru ovog programa testirana je strategija "hromozomske" hijerarhijske podjele genoma, koju su učesnici međunarodnog konzorcijuma koristili za dešifriranje genoma riže. Međutim, ako su u proučavanju ljudskog genoma frakcije pojedinačnih hromozoma izolovane različitim metodama, onda je materijal specifičan za pojedine hromozome riže i njihove pojedinačne regije dobiven laserskom mikrodisekcijom (izrezivanje mikroskopskih objekata). Na stakalcu mikroskopa, gdje se nalaze hromozomi riže, pod utjecajem laserskog snopa sve je izgorjelo, osim hromozoma ili njegovih dijelova predviđenih za analizu. Preostali materijal se koristi za kloniranje i sekvenciranje.
Objavljeni su brojni izvještaji o rezultatima sekvencioniranja pojedinačnih fragmenata genoma riže, provedenih sa visokom preciznošću i detaljnošću, karakterističnom za hijerarhijsku tehnologiju. Vjerovalo se da će utvrđivanje kompletne primarne strukture genoma riže biti završeno do kraja 2003. – sredine 2004. godine, a rezultati će, zajedno sa podacima o primarnoj strukturi genoma Arabidopsis, biti široko korišteni u komparativnom radu. genomika drugih biljaka.
Međutim, početkom 2002. godine dvije istraživačke grupe - jedna iz Kine, druga iz Švicarske i Sjedinjenih Država - objavile su rezultate kompletnog nacrta (približnog) sekvenciranja genoma riže, izvedenog korištenjem tehnologije potpunog kloniranja. Za razliku od etapnog (hijerarhijskog) istraživanja, ukupni pristup se zasniva na istovremenom kloniranju cjelokupne genomske DNK u jednom od virusnih ili bakterijskih vektora i dobivanju značajnog (ogromnog za srednje i velike genome) broja pojedinačnih klonova koji sadrže različite DNK segmenti. Na osnovu analize ovih sekvenciranih sekcija i preklapanja identičnih terminalnih sekcija DNK, formira se kontig - lanac sekvenci DNK spojenih zajedno. Opšti (ukupni) kontig je primarna struktura čitavog genoma, ili barem pojedinačnog hromozoma.
U takvoj šematskoj prezentaciji, strategija totalnog kloniranja izgleda jednostavno. Zapravo, nailazi na ozbiljne poteškoće povezane s potrebom za dobivanjem ogromnog broja klonova (općenito je prihvaćeno da se genom ili njegova regija koja se proučava moraju preklapati klonovima najmanje 10 puta), ogromnom količinom sekvenciranja i izuzetno složenim rad na pristajanju klonova koji zahtijeva učešće stručnjaka za bioinformatiku. Ozbiljna prepreka potpunom kloniranju su različiti repetitivni segmenti DNK, čiji broj, kao što je već spomenuto, naglo raste kako se veličina genoma povećava. Stoga se strategija totalnog sekvenciranja uglavnom koristi u proučavanju genoma virusa i mikroorganizama, iako se uspješno koristi za proučavanje genoma višećelijskog organizma Drosophila.
Rezultati ukupnog sekvenciranja ovog genoma su "superimirani" na ogroman niz informacija o njegovoj hromozomskoj, genskoj i molekularnoj strukturi dobijenih tokom skoro 100-godišnjeg perioda proučavanja Drosophile. Pa ipak, u pogledu stepena sekvenciranja, genom Drosophila (66% ukupne veličine genoma) je značajno inferioran u odnosu na genom Arabidopsis (92%), uprkos njihovim prilično bliskim veličinama - 180 miliona i 125 miliona baznih parova, respektivno. . Stoga je nedavno predloženo da se nazove mješovita tehnologija, koja je korištena za sekvenciranje genoma Drosophila.
Da bi sekvencionirali genom riže, gore spomenute istraživačke grupe su uzele dvije njegove podvrste, koje se najčešće uzgajaju u azijskim zemljama, - Oriza saliva L. ssp indicaj i Oriza saliva L. sspjaponica. Rezultati njihovih studija se u mnogo čemu poklapaju, ali se u mnogo čemu razlikuju. Tako su predstavnici obe grupe izjavili da su dostigli približno 92-93% preklapanja genoma sa kontigom. Pokazalo se da je oko 42% genoma riže predstavljeno kratkim ponavljanjima DNK koji se sastoje od 20 baznih parova, a većina mobilnih DNK elemenata (transposona) nalazi se u intergenskim regijama. Međutim, podaci o veličini genoma riže značajno se razlikuju.
Za japansku podvrstu, veličina genoma je određena na 466 miliona parova baza, a za indijsku 420 miliona. Razlog za ovo odstupanje nije jasan. To može biti posljedica različitih metodoloških pristupa u određivanju veličine nekodirajućeg dijela genoma, odnosno ne odražava pravo stanje stvari. Ali moguće je da postoji razlika od 15% u veličini proučavanih genoma.
Druga velika razlika otkrivena je u broju pronađenih gena: za japansku podvrstu od 46 022 do 55 615 gena po genomu, a za indijsku od 32 000 do 50 000. Razlog ovog odstupanja nije jasan.
Nepotpunost i nedosljednost primljenih informacija konstatuje se u komentarima na objavljene članke. Takođe izražava nadu da će praznine u poznavanju genoma riže biti eliminirane upoređivanjem podataka "grubog sekvenciranja" s rezultatima detaljnog, hijerarhijskog sekvenciranja koje su sproveli učesnici Međunarodnog projekta genoma riže.
KOMPARATIVNA I FUNKCIONALNA BILJNA GENOMIKA
Dobiveni opsežni podaci, od kojih je polovina (rezultati kineske grupe) javno dostupna, nesumnjivo otvaraju široke perspektive kako za proučavanje genoma riže tako i za biljnu genomiku općenito. Poređenje svojstava genoma Arabidopsis i riže pokazalo je da se većina gena (do 80%) identificiranih u genomu Arabidopsis također nalazi u genomu riže; . Istovremeno, 98% gena čija je primarna struktura utvrđena za druge žitarice pronađeno je u genomu pirinča.
Značajna (gotovo dvostruka) razlika između broja gena u riži i Arabidopsisu je zbunjujuća. Istovremeno, podaci nacrta dekodiranja genoma riže dobiveni korištenjem totalnog sekvenciranja praktično se ne upoređuju sa opsežnim rezultatima proučavanja genoma riže metodom hijerarhijskog kloniranja i sekvenciranja, odnosno onoga što je urađeno u na genom Drosophila nije sprovedena. Stoga ostaje nejasno da li razlika u broju gena kod Arabidopsisa i riže odražava pravo stanje stvari ili se objašnjava razlikom u metodološkim pristupima.
Za razliku od genoma Arabidopsis, podaci o genima blizancima u genomu riže nisu dati. Moguće je da njihova relativna količina može biti veća u riži nego u Arabidopsisu. Ovu mogućnost indirektno potkrepljuju podaci o prisutnosti poliploidnih oblika riže. Više jasnoće po ovom pitanju može se očekivati nakon što se završi Međunarodni projekat genoma riže i dobije detaljna slika primarne strukture DNK ovog genoma. Ozbiljne osnove za takvu nadu daje činjenica da se nakon objavljivanja radova o grubom sekvenciranju genoma riže, broj publikacija o strukturi ovog genoma naglo povećao, a posebno su se pojavile informacije o detaljnom sekvenciranju. njegovih 1 i 4 hromozoma.
Poznavanje, barem približno, broja gena u biljkama je od fundamentalnog značaja za uporednu biljnu genomiku. U početku se vjerovalo da, budući da su sve cvjetnice vrlo bliske jedna drugoj po svojim fenotipskim karakteristikama, njihovi genomi bi također trebali biti slični. A ako proučimo genom Arabidopsis, dobićemo informacije o većini genoma drugih biljaka. Indirektna potvrda ove pretpostavke su rezultati sekvenciranja mišjeg genoma, koji je iznenađujuće blizak ljudskom genomu (oko 30 hiljada gena, od kojih se samo 1 tisuću pokazalo drugačijim).
Može se pretpostaviti da razlog razlika između genoma Arabidopsis i riže leži u njihovoj pripadnosti različitim klasama biljaka - dvosupnicama i jednosupnicama. Da bi se ovo pitanje razjasnilo, vrlo je poželjno poznavati barem grubu primarnu strukturu neke druge monokotiledonske biljke. Najrealniji kandidat mogao bi biti kukuruz, čiji je genom približno jednak ljudskom genomu, ali još uvijek mnogo manji od genoma drugih žitarica. Nutritivna vrijednost kukuruza je dobro poznata.
Ogroman materijal dobiven kao rezultat sekvenciranja genoma Arabidopsis i riže postupno postaje osnova za opsežno proučavanje biljnih genoma korištenjem komparativne genomike. Ovakve studije su od opšteg biološkog značaja, jer nam omogućavaju da ustanovimo glavne principe organizacije biljnog genoma u celini i njihovih pojedinačnih hromozoma, da identifikujemo zajedničke karakteristike strukturu gena i njihovih regulatornih regiona, da se uzme u obzir odnos funkcionalno aktivnog (genskog) dela hromozoma i različitih intergenskih DNK regiona koji ne kodiraju proteine. Komparativna genetika također postaje sve važnija za razvoj ljudske funkcionalne genomike. Za komparativne studije provedeno je sekvencioniranje genoma ribe pahuljice i miša.
Jednako važno je proučavanje pojedinačnih gena odgovornih za sintezu pojedinačnih proteina koji određuju specifične tjelesne funkcije. Upravo u otkrivanju, izolaciji, sekvenciranju i određivanju funkcije pojedinačnih gena leži praktičan, prvenstveno medicinski značaj programa Ljudski genom. Ovu okolnost je prije nekoliko godina uočio J. Watson, koji je naglasio da će program Ljudski genom biti završen tek kada se utvrde funkcije svih ljudskih gena.

Rice. četiri. Klasifikacija prema funkciji gena Arabidopsis
1 - geni za rast, diobu i sintezu DNK; 2 - geni za sintezu RNK (transkripcija); 3 - geni za sintezu i modifikaciju proteina; 4 - geni za razvoj, starenje i ćelijsku smrt; 5 - geni ćelijskog metabolizma i energetskog metabolizma; 6 - geni međućelijske interakcije i prijenosa signala; 7 - geni za obezbeđivanje drugih ćelijskih procesa; 8 - geni sa nepoznatom funkcijomŠto se tiče funkcije biljnih gena, znamo manje od jedne desetine onoga što znamo o ljudskim genima. Čak i kod Arabidopsis, čiji genom daleko nadmašuje ljudski genom u smislu stepena proučavanja, funkcija skoro polovine njegovih gena ostaje nepoznata (slika 4). U međuvremenu, pored gena uobičajenih za životinje, biljke imaju značajan broj gena koji su specifični samo (ili barem pretežno) za njih. Radi se o o genima uključenim u transport vode i sintezi stanične stijenke, kojih nema kod životinja, o genima koji osiguravaju stvaranje i funkcioniranje hloroplasta, fotosintezu, fiksaciju dušika i sintezu brojnih aromatičnih proizvoda. Ovaj popis se može nastaviti, ali je već jasno s kojim se teškim zadatkom suočava funkcionalna genomika biljaka.
Puno sekvenciranje genoma daje bliske istinitim informacije o ukupnom broju gena u datom organizmu, omogućava da se manje ili više detaljne i pouzdane informacije o njihovoj strukturi smjeste u banke podataka, te olakšava rad na izolaciji i proučavanju pojedinačnih gena. Međutim, sekvenciranje genoma nikako ne znači uspostavljanje funkcije svih gena.
Jedan od pristupa funkcionalne genomike koji najviše obećava zasniva se na identifikaciji radnih gena koji se koriste za transkripciju (čitanje) mRNA. Ovaj pristup, uključujući moderna tehnologija mikromreža, omogućava vam da istovremeno identifikujete do desetine hiljada funkcionalnih gena. Nedavno je istraživanje biljnih genoma počelo korištenjem ovog pristupa. Za Arabidopsis je bilo moguće dobiti oko 26 hiljada pojedinačnih transkripata, što uvelike olakšava mogućnost određivanja funkcije gotovo svih njegovih gena. U krompiru je bilo moguće identifikovati oko 20.000 radnih gena koji su važni za razumevanje kako procesa rasta i formiranja gomolja, tako i procesa bolesti krompira. Pretpostavlja se da će ovo saznanje povećati otpornost jednog od najvažnijih prehrambenih proizvoda na patogene.
Logičan razvoj funkcionalne genomike bila je proteomika. Ovo novo polje nauke proučava proteom, koji se obično shvata kao kompletan skup proteina u ćeliji u određenom trenutku. Takav skup proteina, koji odražava funkcionalno stanje genoma, stalno se mijenja, dok genom ostaje nepromijenjen.
Proučavanje proteina se dugo koristi za procjenu aktivnosti biljnih genoma. Kao što je poznato, enzimi prisutni u svim biljkama razlikuju se kod pojedinačnih vrsta i varijanti u redoslijedu aminokiselina. Takvi enzimi, sa istom funkcijom, ali različitim slijedom pojedinačnih aminokiselina, nazivaju se izoenzimi. Imaju različita fizičko-hemijska i imunološka svojstva (molekularna težina, naboj), koja se mogu otkriti hromatografijom ili elektroforezom. Dugi niz godina ove metode se uspješno koriste za proučavanje takozvanog genetskog polimorfizma, odnosno razlika između organizama, sorti, populacija, vrsta, posebno pšenice i srodnih oblika žitarica. Nedavno je, međutim, zbog brzog razvoja metoda analize DNK, uključujući sekvenciranje, proučavanje polimorfizma proteina zamijenjeno proučavanjem polimorfizma DNK. Međutim, direktno proučavanje spektra skladišnih proteina (prolamina, glijadina, itd.), koji određuju glavne nutritivne osobine žitarica, ostaje važna i pouzdana metoda za genetičku analizu, selekciju i proizvodnju sjemena poljoprivrednih biljaka.
Poznavanje gena, mehanizama njihove ekspresije i regulacije izuzetno je važno za razvoj biotehnologije i proizvodnju transgenih biljaka. Poznato je da impresivni uspjesi u ovoj oblasti izazivaju dvosmislenu reakciju ekološke i medicinske zajednice. Međutim, postoji područje biljne biotehnologije gdje se ovi strahovi, ako ne i potpuno neutemeljeni, onda, u svakom slučaju, čine malo važnima. Riječ je o stvaranju transgenih industrijskih biljaka koje se ne koriste kao prehrambeni proizvodi. Indija je nedavno pobrala prvi usev transgenog pamuka koji je otporan na brojne bolesti. Postoje informacije o uvođenju posebnih gena koji kodiraju pigmentne proteine u genom pamuka i proizvodnji pamučnih vlakana koja ne zahtijevaju umjetno bojenje. Još jedna industrijska kultura koja može biti predmet efikasnog genetskog inženjeringa je lan. Nedavno se raspravljalo o njegovoj upotrebi kao alternativi pamuku za tekstilne sirovine. Ovaj problem je izuzetno važan za našu zemlju, koja je izgubila sopstvene izvore sirovog pamuka.
PERSPEKTIVE ZA PROUČAVANJE BILJNIH GENOMA
Očigledno je da će se strukturne studije biljnih genoma zasnivati na pristupima i metodama komparativne genomike, koristeći rezultate dešifriranja genoma Arabidopsis i riže kao glavnog materijala. Bitnu ulogu u razvoju komparativne biljne genomike nesumnjivo će imati informacije koje će prije ili kasnije dobiti totalno (grubo) sekvenciranje genoma drugih biljaka. U ovom slučaju, komparativna genomika biljaka će se zasnivati na uspostavljanju genetskih odnosa između pojedinačnih lokusa i hromozoma koji pripadaju različitim genomima. Nećemo se fokusirati toliko na opštu genomiku biljaka koliko na selektivnu genomiku pojedinačnih hromozomskih lokusa. Na primjer, nedavno je pokazano da se gen odgovoran za jarovizaciju nalazi na VRn-AI lokusu heksaploidnog hromozoma 5A pšenice i Hd-6 lokusu rižinog hromozoma 3.
Razvoj ovih studija bit će snažan poticaj za identifikaciju, izolaciju i sekvenciranje mnogih funkcionalno važnih biljnih gena, posebno gena odgovornih za otpornost na bolesti, otpornost na sušu i prilagodljivost različitim uvjetima uzgoja. Sve više će se koristiti funkcionalna genomika, zasnovana na masovnoj detekciji (skriningu) gena koji funkcionišu u biljkama.
Možemo predvidjeti dalje unapređenje hromozomskih tehnologija, prvenstveno metode mikrodisekcije. Njegova upotreba dramatično proširuje mogućnosti genomskog istraživanja bez velikih troškova, kao što je, na primjer, sekvenciranje ukupnog genoma. Metoda lokalizacije na hromozomima biljaka pojedinih gena uz pomoć hibridizacije će se dalje širiti. in situ. Trenutno je njegova primjena ograničena ogromnim brojem ponavljajućih sekvenci u biljnom genomu, a moguće i posebnostima strukturne organizacije biljnih hromozoma.
Kromosomske tehnologije će u doglednoj budućnosti postati od velike važnosti za evolucijsku genomiku biljaka. Ove relativno jeftine tehnologije omogućavaju brzu procjenu intra- i interspecifične varijabilnosti, proučavanje složenih alopoliploidnih genoma tetraploidne i heksaploidne pšenice, tritikala; analizirati evolucijske procese na hromozomskom nivou; istražuju formiranje sintetičkih genoma i unošenje (introgresija) stranog genetskog materijala; identificirati genetske veze između pojedinačnih hromozoma razne vrste.
Za karakterizaciju genoma koristit će se proučavanje kariotipa biljaka klasičnim citogenetskim metodama, obogaćenim molekularno-biološkom analizom i kompjuterskom tehnologijom. Ovo je posebno važno za proučavanje stabilnosti i varijabilnosti kariotipa na nivou ne samo pojedinačnih organizama, već i populacija, varijeteta i vrsta. Konačno, teško je zamisliti kako se broj i spektri kromosomskih preuređivanja (aberacije, mostovi) mogu procijeniti bez upotrebe diferencijalnih metoda bojenja. Ovakve studije su izuzetno obećavajuće za praćenje životne sredine prema stanju biljnog genoma.
AT moderna Rusija direktno sekvenciranje biljnih genoma je malo vjerovatno. Takav posao, koji iziskuje velika ulaganja, nepodnošljiv je za našu sadašnju ekonomiju. U međuvremenu, podaci o strukturi genoma Arabidopsis i riže, dobijeni od strane svjetske nauke i dostupni u međunarodnim bankama podataka, dovoljni su za razvoj domaće biljne genomike. Može se predvidjeti proširenje proučavanja biljnih genoma zasnovanih na komparativnim genomskim pristupima za rješavanje specifičnih problema oplemenjivanja i uzgoja biljaka, kao i za proučavanje porijekla različitih biljnih vrsta od velikog ekonomskog značaja.
Može se pretpostaviti da će genomski pristupi kao što je genetska tipizacija (RELF, RAPD, AFLP analize, itd.), koji su prilično pristupačni za naš budžet, imati široku primjenu u domaćoj oplemenjivačkoj praksi i biljnoj proizvodnji. Paralelno sa direktnim metodama za određivanje polimorfizma DNK, pristupi zasnovani na proučavanju polimorfizma proteina, prvenstveno proteina skladištenja žitarica, koristiće se za rješavanje problema genetike i oplemenjivanja biljaka. Kromosomske tehnologije će se široko koristiti. Relativno su jeftini, njihov razvoj zahtijeva prilično umjerena ulaganja. U oblasti proučavanja hromozoma domaća nauka nije inferiorna u odnosu na svet.
Treba naglasiti da je naša nauka dala značajan doprinos nastanku i razvoju biljne genomike [ , ].
Temeljnu ulogu odigrao je N.I. Vavilov (1887-1943).
U molekularnoj biologiji i biljnoj genomici, pionirski doprinos A.N. Belozerski (1905-1972).
U oblasti hromozomskih studija potrebno je istaći rad izuzetnog genetičara S.G. Navašin (1857-1930), koji je prvi otkrio satelitske hromozome u biljkama i dokazao da je moguće razlikovati pojedinačne hromozome prema karakteristikama njihove morfologije.
Još jedan klasik Ruska nauka G.A. Levitsky (1878-1942) je detaljno opisao hromozome raži, pšenice, ječma, graška i šećerne repe, uveo pojam "kariotip" u nauku i razvio doktrinu o tome.
Savremeni stručnjaci, oslanjajući se na dostignuća svjetske nauke, mogu dati značajan doprinos daljem razvoju biljne genetike i genomike.
Autor se od srca zahvaljuje akademiku Yu.P. Altukhovu za kritičku raspravu o članku i vrijedne savjete.Rad tima na čelu sa autorom članka podržala je Ruska fondacija za osnovna istraživanja (grantovi br. 99-04-48832; 00-04-49036; 00-04-81086), Ruska Federacija za podršku naučnim školama (grantovi br. 00-115-97833 i NSh-1794.2003.4) i Program Ruska akademija nauke "Molekularno genetski i hromozomski markeri u razvoju savremenih metoda oplemenjivanja i proizvodnje sjemena".
LITERATURA
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Uvod u biljnu genomiku // Molekularna biologija. 2001. V. 35. S. 339-348. 2. Pen E. Bonanza za biljnu genomiku // Science. 1998. V. 282. P. 652-654. 3. Biljna genomika, Proc. Natl. Akad. sci. SAD. 1998. V. 95. P. 1962-2032. 4. Cartel N.A. i sl. Genetika. Encyclopedic Dictionary. Minsk: Tehnologija, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferencijacija genoma u Aegilopsu. 1. Distribucija visoko repetitivnih sekvenci DNK na hromozomima diploidnih vrsta, Genom. 1996. V. 39. P. 293-306. Povijest analize hromozoma // Biol. membrane. 2001. T. 18. S. 164-172.
Dana 05.09.2011. u 09:36, Limarev je rekao:
Limarev V.N.
Dešifrovanje ljudskog genoma.
Fragment iz knjige L.G. Pučko: "Radietsko znanje o čovjeku"
Za rješavanje problema dešifriranja genoma organiziran je međunarodni projekat "Ljudski genom" sa budžetom od milijardu dolara.
Do 2000. godine mapa ljudskog genoma je bila praktički kompletna. Geni su prebrojani, identificirani i zabilježeni u bazama podataka. To su ogromne količine informacija.
Za snimanje ljudskog genoma u digitalizovanom obliku potrebno je oko 300 terabajta računarske memorije, što je ekvivalentno 3.000 čvrstih diskova kapaciteta 100 gigabajta.
Ispostavilo se. Da osoba nema stotine hiljada, kako se ranije mislilo, već nešto više od 30 hiljada gena. Muva ima Drosophila, ima ih samo upola manje - oko 13 hiljada, a miš ima skoro isti broj kao i osoba. Geni jedinstveni za ljude u dekodiranom genomu su samo oko 1%. Većinu spirale DNK, kako se ispostavilo, ne zauzimaju geni, već takozvani "prazni dijelovi", u kojima geni jednostavno nisu kodirani, kao i dvostruki fragmenti koji se ponavljaju jedan za drugim, značenje i značenje od kojih je nejasno.
Jednom riječju, ispostavilo se da geni nisu čak ni cigle života, već samo elementi plana, prema kojem se gradi izgradnja organizma. Cigle, kao što se vjerovalo prije vrhunca genetike, su proteini.
Postalo je apsolutno očito da se u 1% gena jedinstvenih za ljude ne može kodirati tako ogromna količina informacija koja razlikuje ljude od miševa. Gdje su sve informacije pohranjene? Za mnoge naučnike postaje nesumnjiva činjenica da je bez Božanskog principa nemoguće objasniti prirodu čoveka. Brojni naučnici sugerišu da je, u okviru postojećih ideja o ljudskom tijelu, u osnovi nemoguće dešifrirati ljudski genom.
Svijet se ne poznaje - može se spoznati (moji komentari na članak).
1) Razmotrite fragment: “Bez božanskog principa nemoguće je objasniti prirodu čovjeka.”
Gore navedene informacije ne govore ništa o tome.
Genom, zaista, ima složeniju strukturu nego što se ranije mislilo.
Ali, uostalom, kompjuter spomenut u članku ne sastoji se samo od memorijskih ćelija.
Računar ima dvije memorije: dugoročnu i operativnu, kao i procesor u kojem se obrađuju informacije. Učestvuje u obradi informacija i elektromagnetnog polja. Da bi se dešifrovale informacije o genomu, potrebno je razumjeti kako se to događa, ne samo skladištenje informacija, već i njihova obrada. Prihvaćam i ideju da se dio informacija pohranjuje snimljeno od strane elektromagnetno polje. I takođe izvan osobe, kao što sam već napisao, u posebnim informacionim centrima Višeg uma.
Zamislite neprekidni tekst kodiran u binarnom kodu 0 ili 1 Morzeove azbuke, dok ne znate na kom jeziku (engleskom ili francuskom....) je napisan, a ne znate da se ovaj kontinuirani tekst sastoji od riječi, rečenica , paragrafi, poglavlja, tomovi, police, ormarići, itd.
U biologiji je skoro isto, samo što je ovde sve kodirano četvorocifrenim kodom i mi smo do sada dešifrovali redosled elementarnih gena + - / *, ali ne znamo jezik i shodno tome riječi, rečenice, paragrafi, poglavlja, tomovi, police, ormari, itd. Dešifrovani genom za nas je i dalje čvrst tekst šifre od 4 žitarice i gotovo je nemoguće sve to proučiti direktno.
Ali ispostavilo se da se u određenim vremenskim periodima (kako u pojedincu i njegovoj kohorti generacija tako i u vrsti, rodu) neki geni i njihovi kompleksi (odgovorni za riječi, rečenice, pasuse, poglavlja, tomove, police, ormariće, itd. itd.) su aktivni, au drugim periodima evolucije pasivni, što sam posredno odredio raznim poligenskim osobinama (što je prikazano u temi Univerzalni periodični zakon evolucije).
Do sada postoje samo dvije metode za proučavanje gena, ovo je jednostavan laboratorijski proračun sume gena (DNK) u uzorku i postoji uređaj koji broji količinu proizvedenih RNA proteina zaglavio na generisanom elektronskom čipu specifična DNK, ali pošto je ogromna količina DNK aktivna u svakom trenutku i, shodno tome, ogromna količina različitih proteina se proizvodi kroz RNK, vrlo je teško odvojiti "ove rezance kašikom, vilicom i japanskim štapićima" u ovoj supi i pronađite ono što tražite - da pronađete uzročno-posledične veze između određene DNK (kao DNK kompleksa) i njenog utjecaja na poligensku osobinu.
Čini se da sam pronašao jednostavnu metodu kako da sredim cijelu ovu juhu od DNK, RNK i njihovih proteina koji određuju stepen poligene osobine.
Kako se ispostavilo, svaka poligenska osobina u evolutivnom redu jedinke (kohorta generacija, vrsta i rod) je periodična, stoga moraju biti periodična u RNA i DNK aktivnosti, te je stoga potrebno samo pronaći (prvo zalazeći u genetske detalje) korelaciju između metričke promjene u poligenskoj osobini (kod pojedinca, grupe generacija, vrste, roda...) i proporcionalne tim periodima, odgovarajuće aktivnosti RNK, DNK.
) nalazi se u genomu voćne mušice ( Drosophila ananassae) kompletna kopija bakterijskog genoma parazita Wolbachia.
Bakterija Wolbachia živi u citoplazmi ćelija domaćina i poznata je po tome što je naučila fino regulirati reprodukciju, razvoj, pa čak i evoluciju svojih domaćina. Stoga ga često nazivaju „mikrobnim manipulatorom“ ili „gospodarom muva“ (pošto živi u ćelijama insekata).
Studija je započela kada je Julie Dunning-Hotopp iz JCVI otkrila kako određeni geni Wolbachia "sarađuju" sa genima Drosophila kao da su dio istog genoma.
Michael Clark - istraživač na Univerzitetu u Rochesteru - naselio je koloniju Drosophila ananassae u laboratoriju da otkrije tajnu sa Warrenom.
Gen Wolbachia u genomu Drosophila (ilustrovan na Univerzitetu u Rochesteru).
„Nekoliko meseci sam mislio da sam u nečemu pogrešio“, kaže Klark, „čak sam sugerisao da se razvila rezistencija na antibiotike, jer sam pronalazio svaki gen Wolbachije iznova i iznova. Kada sam konačno uzeo maramice koje sam ostavio na miru prije nekoliko mjeseci, nisam našao samu Wolbachiu.
Sada Warren i Clarke pokušavaju shvatiti koja je prednost umetanja tako velikog dijela DNK za Drosophilu - možda "strani" geni pružaju domaćinu neke nove mogućnosti.

I tako geni Wolbachia prelaze u DNK domaćina (ilustracija Nicolle Rager Fuller, National Science).
Rezultati studije objavljeni su u članku u časopisu Science. U njemu autori sugeriraju da se horizontalni prijenos gena (prijenos gena između nepovezanih vrsta) događa između bakterija i višećelijskih organizama u našem svijetu mnogo češće nego što se mislilo.
Dešifrovanje molekularno genetskih mehanizama manipulacija koje provodi Wolbachia sa svojim domaćinima dat će čovjeku moćna nova sredstva utjecaja na žive organizme i prirodu u cjelini.
Međutim, nisu svi insekti podložni lošem utjecaju Wolbachia. Na primjer, leptiri sa Samoanskih ostrva su "naučili" da štite svoje mužjake. Pitam se da li će malarični komarci, koje žele da zaraze ovom bakterijom, naučiti da se bore protiv nje?
Povodom 50. godišnjice otkrića strukture DNK
A.V. Zelenin
BILJNI GENOM
A. V. Zelenin
Zelenin Aleksandar Vladimirovič- d.b.n.,
šef laboratorije Instituta za molekularnu biologiju. V.A. Engelhardt RAS.
Impresivna dostignuća programa "Ljudski genom", kao i uspeh rada na dešifrovanju takozvanih ekstra malih (virusi), malih (bakterije, kvasac) i srednjih (okrugli crvi, Drosophila) genoma, omogućili su premještanje na veliku studiju velikih i ekstra velikih biljnih genoma. Hitna potreba za detaljnim proučavanjem genoma ekonomski najvažnijih biljaka naglašena je na skupu o biljnoj genomici održanom 1997. godine u Sjedinjenim Državama [ , ]. Tokom godina koje su prošle od tog vremena, postignuti su nesumnjivi uspjesi u ovoj oblasti. 2000. godine pojavila se publikacija o potpunom sekvenciranju (određivanje linearne sekvence nukleotida cjelokupne nuklearne DNK) genoma male gorušice - Arabidopsis, 2001. godine - o preliminarnom (nacrtu) sekvenciranju genoma riže. Radovi na sekvenciranju genoma velikih i super velikih biljaka (kukuruz, raž, pšenica) su više puta izvještavani, međutim, ti izvještaji nisu sadržavali konkretne informacije i više su bili po prirodi deklaracija namjere.
Pretpostavlja se da će dekodiranje biljnih genoma otvoriti široke perspektive za nauku i praksu. Prije svega, identifikacija novih gena i lanac njihove genetske regulacije značajno će povećati produktivnost biljaka korištenjem biotehnoloških pristupa. Otkrivanjem, izolacijom, reprodukcijom (kloniranjem) i sekvenciranjem gena odgovornih za tako važne funkcije biljnog organizma kao što su reprodukcija i produktivnost, procesi varijabilnosti, otpornost na štetne faktore okoline, kao i homologno uparivanje hromozoma, dolazi do pojave povezane su nove mogućnosti za unapređenje procesa uzgoja. Konačno, izolovani i klonirani geni se mogu koristiti za dobijanje transgenih biljaka sa fundamentalno novim svojstvima i za analizu mehanizama regulacije aktivnosti gena.
Važnost proučavanja biljnih genoma naglašava i činjenica da je do sada broj lokaliziranih, kloniranih i sekvenciranih biljnih gena mali i varira, prema različitim procjenama, između 800 i 1200. To je 10-15 puta manje nego za na primjer, kod ljudi.
Sjedinjene Američke Države ostaju nesumnjivi lider u istraživanju genoma biljaka velikih razmjera, iako se intenzivne studije genoma riže provode u Japanu, a posljednjih godina iu Kini. U dešifrovanju genoma Arabidopsis, pored američkih laboratorija, aktivno su učestvovale i evropske istraživačke grupe. Očigledno vodstvo Sjedinjenih Država izaziva ozbiljnu zabrinutost evropskih naučnika, što su jasno izrazili na skupu pod značajnim naslovom "Izgledi za genomiku u postgenomskoj eri", održanom krajem 2000. godine u Francuskoj. Napredak američke nauke u proučavanju genoma poljoprivrednih biljaka i stvaranju transgenih biljnih formi, prema evropskim naučnicima, prijeti da će u ne tako dalekoj budućnosti (dvije do pet decenija), kada će porast populacije staviti čovječanstvo u lice opšteg prehrambena kriza, evropska ekonomija i nauka će postati zavisni od američke tehnologije. S tim u vezi, najavljeno je stvaranje francusko-njemačkog naučnog programa za proučavanje biljnih genoma ("Plantgene") iu njega su uložena značajna ulaganja.
Očigledno je da bi problemi biljne genomike trebali privući veliku pažnju ruskih naučnika i organizatora nauke, ali i vlasti, jer se ne radi samo o naučnom prestižu, već i o nacionalnoj sigurnosti zemlje. Za deceniju ili dve, hrana će postati najvažniji strateški resurs.
TEŠKOĆE U PROUČAVANJU BILJNIH GENOMA
Proučavanje genoma biljaka mnogo je teži zadatak od proučavanja genoma ljudi i drugih životinja. To je zbog sljedećih okolnosti:
ogromne veličine genoma, koje dosežu desetke, pa čak i stotine milijardi baznih parova (bp) za pojedinačne biljne vrste: genomi glavnih ekonomski važnih biljaka (osim riže, lana i pamuka) su ili blizu ljudskog genoma ili su veći od to više puta (tabela);HROMOSOMSKA STUDIJA GENOMAOštre fluktuacije u broju hromozoma u različitim biljkama - od dva kod nekih vrsta do nekoliko stotina u drugih, i nije moguće utvrditi strogu korelaciju između veličine genoma i broja hromozoma;
Obilje poliploida (koji sadrže više od dva genoma po ćeliji) formira se sa sličnim, ali ne identičnim genomima (alpoliploidija);
Ekstremno obogaćivanje biljnih genoma (do 99%) "beznačajne" (nekodirajuće, odnosno ne sadrži gene) DNK, što otežava spajanje sekvencioniranih fragmenata (poređanje u ispravnom redoslijedu) u zajednički DNK region velike veličine (kontig);
Nepotpuno (u poređenju sa genomima drozofile, čoveka i miša) morfološko, genetsko i fizičko mapiranje hromozoma;
Praktična nemogućnost izolacije pojedinačnih hromozoma u čistom obliku korišćenjem metoda koje se obično koriste u tu svrhu za ljudske i životinjske hromozome (razvrstavanje u struji i upotreba ćelijskih hibrida);
Poteškoća hromozomskog mapiranja (određivanje lokacije na hromozomu) pojedinačnih gena pomoću hibridizacije in situ, kako zbog visokog sadržaja "beznačajne" DNK u biljnim genomima, tako i zbog posebnosti strukturne organizacije biljnih hromozoma;
Evolucijska udaljenost biljaka od životinja, što ozbiljno otežava korištenje informacija dobivenih sekvenciranjem genoma ljudi i drugih životinja za proučavanje genoma biljaka;
Dug proces reprodukcije većine biljaka, što značajno usporava njihovu genetsku analizu.
Kromosomske (citogenetske) studije genoma općenito, a posebno biljaka, imaju dugu povijest. Termin "genom" predložen je za označavanje haploidnog (jednostrukog) skupa hromozoma sa genima koji se nalaze u njima u prvoj četvrtini 20. veka, odnosno mnogo pre uspostavljanja uloge DNK kao nosioca genetske informacije. .
Opis genoma novog, ranije genetski neistraženog višećelijskog organizma obično počinje proučavanjem i opisom kompletnog skupa njegovih hromozoma (kariotipa). To se, naravno, odnosi i na biljke, od kojih veliki broj nije ni počeo da se proučava.
Već na početku istraživanja hromozoma upoređivani su genomi srodnih biljnih vrsta na osnovu analize mejotičke konjugacije (kombinacije homolognih hromozoma) u interspecifičnim hibridima. Tokom proteklih 100 godina, mogućnosti analize hromozoma su se dramatično proširile. Sada se za karakterizaciju biljnih genoma koriste naprednije tehnologije: razne varijante takozvanog diferencijalnog bojenja, koje omogućava identifikaciju pojedinačnih hromozoma prema morfološkim karakteristikama; hibridizacija in situ omogućavajući lokalizaciju specifičnih gena na hromozomima; biohemijske studije ćelijskih proteina (elektroforeza i imunohemija) i, konačno, set metoda zasnovanih na analizi hromozomske DNK do njenog sekvenciranja.

Rice. jedan. Kariotipovi žitarica a - raž (14 hromozoma), b - durum pšenica (28 hromozoma), c - meka pšenica (42 hromozoma), d - ječam (14 hromozoma)Dugi niz godina proučavaju se kariotipovi žitarica, prvenstveno pšenice i raži. Zanimljivo je da je kod različitih vrsta ovih biljaka broj hromozoma različit, ali uvijek višestruk od sedam. Pojedine vrste žitarica mogu se pouzdano prepoznati po kariotipu. Na primjer, genom raži sastoji se od sedam parova velikih hromozoma sa intenzivno obojenim heterokromatskim blokovima na njihovim krajevima, koji se često nazivaju segmenti ili trake (slika 1a). Genomi pšenice već imaju 14 i 21 par hromozoma (sl. 1, b, c), a distribucija heterokromatskih blokova u njima nije ista kao u hromozomima raži. Pojedinačni genomi pšenice, označeni A, B i D, također se međusobno razlikuju. Povećanje broja hromozoma sa 14 na 21 dovodi do nagle promjene svojstava pšenice, što se ogleda u njihovim nazivima: durum, ili testenina, pšenica i mekana, ili hleb, pšenica. D gen, koji sadrži gene za proteine glutena, koji daje testu takozvanu klijavost, odgovoran je za sticanje visokih pekarskih svojstava mekom pšenicom. Upravo ovom genomu se pridaje posebna pažnja u selekcijskom poboljšanju hljebne pšenice. Druga žitarica sa 14 hromozoma, ječam (slika 1, d), obično se ne koristi za pravljenje hleba, ali je glavna sirovina za proizvodnju uobičajenih proizvoda kao što su pivo i viski.
Intenzivno se istražuju hromozomi nekih samoniklih biljaka koji se koriste za poboljšanje kvaliteta najvažnijih poljoprivrednih vrsta, kao što je divlji srodnik pšenice - Aegilops. Nove biljne forme nastaju ukrštanjem (slika 2) i selekcijom. Posljednjih godina značajno poboljšanje u istraživačkim metodama omogućilo je početak proučavanja biljnih genoma, čije su karakteristike kariotipova (uglavnom mala veličina kromosoma) činile ranije nedostupnim za kromosomsku analizu. Dakle, tek nedavno su prvi put identificirani svi hromozomi pamuka, kamilice i lana.

Rice. 2. Kariotipovi pšenice i hibrid pšenice sa Aegilopsom
a - heksaploidna meka pšenica ( Triticum astivum), koji se sastoji od A, B i O genoma; b - tetraploidna pšenica ( Triticum timopheevi), koji se sastoji od A i G genoma. sadrži gene za otpornost na većinu bolesti pšenice; c - hibridi Triticum astivum X Triticum timopheevi otporan na pepelnicu i rđu, jasno je vidljiva zamjena dijela hromozomaPRIMARNA STRUKTURA DNK
Razvojem molekularne genetike proširio se i sam koncept genoma. Sada se ovaj pojam tumači i u klasičnom hromozomskom i u modernom molekularnom smislu: cjelokupni genetski materijal pojedinačnog virusa, ćelije i organizma. Naravno, nakon proučavanja kompletne primarne strukture genoma (kako se često naziva kompletan linearni niz baza nukleinskih kiselina) brojnih mikroorganizama i ljudi, postavilo se pitanje sekvenciranja biljnog genoma.
Od brojnih biljnih organizama, dva su odabrana za proučavanje - Arabidopsis, koji predstavlja klasu dvosupnica (veličina genoma 125 miliona bp), i pirinač iz klase jednosupnica (420-470 miliona bp). Ovi genomi su mali u poređenju sa drugim biljnim genomima i sadrže relativno malo segmenata DNK koji se ponavljaju. Takve karakteristike su davale nadu da će odabrani genomi biti dostupni za relativno brzo određivanje njihove primarne strukture.

Rice. 3. Arabidopsis - mala gorušica - mala biljka iz porodice krstaša ( Brassicaceae). Na prostoru jednakoj površini jednoj stranici našeg časopisa, možete uzgajati do hiljadu pojedinačnih organizama Arabidopsis.Razlog odabira Arabidopsisa nije bila samo mala veličina njegovog genoma, već i mala veličina organizma, što ga čini lakim za uzgoj u laboratoriji (slika 3). Uzeli smo u obzir njegov kratak reproduktivni ciklus, koji omogućava brzo provođenje eksperimenata ukrštanja i selekcije, detaljno proučavanu genetiku, lakoću manipulacije s promjenjivim uvjetima uzgoja (promjena sastava soli u tlu, dodavanje različitih hranjivih tvari, itd.) i ispitivanje djelovanja na biljke raznih mutagenih faktora i patogena (virusa, bakterija, gljivica). Arabidopsis nema ekonomsku vrijednost, stoga je njen genom, zajedno sa genomom miša, nazvan referentnim, ili, manje tačno, modelom.*
* Pojava pojma "model genoma" u ruskoj literaturi rezultat je netačnog prijevoda engleske fraze model genoma. Riječ "model" znači ne samo pridjev "model", već i imenicu "uzorak", "standard", "model". Ispravnije bi bilo govoriti o uzorku genoma ili referentnom genomu.Intenzivan rad na sekvenciranju genoma Arabidopsis započeo je 1996. godine od strane međunarodnog konzorcijuma koji je uključivao naučne institucije i istraživačke grupe iz SAD-a, Japana, Belgije, Italije, Velike Britanije i Njemačke. U decembru 2000. godine postale su dostupne opsežne informacije koje sumiraju određivanje primarne strukture genoma Arabidopsis. Za sekvenciranje je korištena klasična ili hijerarhijska tehnologija: prvo su proučavani pojedinačni mali dijelovi genoma od kojih su sastavljeni veći dijelovi (kontigi), a u završnoj fazi struktura pojedinačnih hromozoma. Nuklearna DNK genoma Arabidopsis raspoređena je na pet hromozoma. Godine 1999. objavljeni su rezultati sekvenciranja dva hromozoma, a pojavljivanje u štampi informacija o primarnoj strukturi preostala tri dovršilo je sekvenciranje cijelog genoma.
Od 125 miliona baznih parova utvrđena je primarna struktura od 119 miliona, što je 92% ukupnog genoma. Ispostavilo se da je samo 8% genoma Arabidopsis koji sadrži velike blokove repetitivnih segmenata DNK nepristupačno za proučavanje. U pogledu kompletnosti i temeljitosti sekvenciranja eukariotskog genoma, Arabidopsis ostaje među prva tri šampiona zajedno sa jednoćelijskim kvascem. Saccharomyces cerevisiae i višećelijski organizam Caenorhabditis elegance(vidi tabelu).
Oko 15.000 pojedinačnih gena koji kodiraju proteine pronađeno je u genomu Arabidopsis. Otprilike 12 000 njih sadržano je kao dvije kopije po haploidnom (jednom) genomu, tako da je ukupan broj gena 27 000. Broj gena u Arabidopsisu se ne razlikuje mnogo od broja gena u organizmima kao što su ljudi i miševi, ali veličina njegovog genoma 25-30 puta manja. Ova okolnost povezana je sa važnim karakteristikama u strukturi pojedinačnih gena Arabidopsis i cjelokupnoj strukturi njenog genoma.
Geni Arabidopsis su kompaktni, sadrže samo nekoliko egzona (protein-kodirajućih regija) razdvojenih kratkim (oko 250 bp) nekodirajućim DNK segmentima (intronima). Intervali između pojedinačnih gena su u prosjeku 4600 parova baza. Poređenja radi, ističemo da ljudski geni sadrže mnogo desetina, pa čak i stotina egzona i introna, a međugenske regije imaju veličine od 10 hiljada parova baza ili više. Pretpostavlja se da je prisustvo malog kompaktnog genoma doprinijelo evolucijskoj stabilnosti Arabidopsis, budući da je njegova DNK u manjoj mjeri postala meta raznih štetnih agenasa, posebno za uvođenje repetitivnih fragmenata DNK (transpozona) sličnih virusu. u genom.
Od ostalih molekularnih karakteristika genoma Arabidopsis, treba napomenuti da su egzoni obogaćeni gvaninom i citozinom (44% u egzonima i 32% u intronima) u poređenju sa životinjskim genima, kao i prisustvo dvostruko ponovljenih (dupliciranih) gena. . Pretpostavlja se da je do takvog udvostručavanja došlo kao rezultat četiri istovremena događaja, koji se sastoje od udvostručavanja (ponavljanja) dijela gena Arabidopsis, odnosno fuzije srodnih genoma. Ovi događaji, koji su se odigrali prije 100-200 miliona godina, manifestacija su općeg trenda poliploidizacije (višestruko povećanje broja genoma u organizmu), koji je karakterističan za biljne genome. Međutim, neke činjenice pokazuju da duplicirani geni u Arabidopsisu nisu identični i funkcioniraju drugačije, što može biti povezano s mutacijama u njihovim regulatornim regijama.
Riža je postala još jedan predmet kompletnog sekvenciranja DNK. Genom ove biljke je također mali (12 hromozoma, što daje ukupno 420-470 miliona bp), samo 3,5 puta veći od genoma Arabidopsis. Međutim, za razliku od Arabidopsisa, riža je od velike ekonomske važnosti, budući da je osnova ishrane više od polovine čovječanstva, dakle, ne samo milijarde potrošača, već i višemilionska armija ljudi aktivno uključenih u veoma naporan proces njenog kultivacije su od vitalnog značaja za poboljšanje svojih svojstava.
Neki istraživači su počeli proučavati genom riže još 1980-ih, ali su te studije dostigle ozbiljne razmjere tek 1990-ih. Godine 1991. u Japanu je kreiran program za dešifrovanje strukture genoma riže, objedinjujući napore mnogih istraživačkih grupa. Na osnovu ovog programa 1997. godine organizovan je Međunarodni projekat genoma riže. Njegovi učesnici odlučili su da koncentrišu svoje napore na sekvencioniranje jedne od podvrsta riže ( Oriza sativajaponica), u čijem proučavanju je do tada već postignut značajan napredak. Ozbiljan podsticaj i, slikovito rečeno, zvijezda vodilja za takav rad bio je program "Ljudski genom".
U okviru ovog programa testirana je strategija "hromozomske" hijerarhijske podjele genoma, koju su učesnici međunarodnog konzorcijuma koristili za dešifriranje genoma riže. Međutim, ako su u proučavanju ljudskog genoma frakcije pojedinačnih hromozoma izolovane različitim metodama, onda je materijal specifičan za pojedine hromozome riže i njihove pojedinačne regije dobiven laserskom mikrodisekcijom (izrezivanje mikroskopskih objekata). Na stakalcu mikroskopa, gdje se nalaze hromozomi riže, pod utjecajem laserskog snopa sve je izgorjelo, osim hromozoma ili njegovih dijelova predviđenih za analizu. Preostali materijal se koristi za kloniranje i sekvenciranje.
Objavljeni su brojni izvještaji o rezultatima sekvencioniranja pojedinačnih fragmenata genoma riže, provedenih sa visokom preciznošću i detaljnošću, karakterističnom za hijerarhijsku tehnologiju. Vjerovalo se da će utvrđivanje kompletne primarne strukture genoma riže biti završeno do kraja 2003. – sredine 2004. godine, a rezultati će, zajedno sa podacima o primarnoj strukturi genoma Arabidopsis, biti široko korišteni u komparativnom radu. genomika drugih biljaka.
Međutim, početkom 2002. godine dvije istraživačke grupe - jedna iz Kine, druga iz Švicarske i Sjedinjenih Država - objavile su rezultate kompletnog nacrta (približnog) sekvenciranja genoma riže, izvedenog korištenjem tehnologije potpunog kloniranja. Za razliku od etapnog (hijerarhijskog) istraživanja, ukupni pristup se zasniva na istovremenom kloniranju cjelokupne genomske DNK u jednom od virusnih ili bakterijskih vektora i dobivanju značajnog (ogromnog za srednje i velike genome) broja pojedinačnih klonova koji sadrže različite DNK segmenti. Na osnovu analize ovih sekvenciranih sekcija i preklapanja identičnih terminalnih sekcija DNK, formira se kontig - lanac sekvenci DNK spojenih zajedno. Opšti (ukupni) kontig je primarna struktura čitavog genoma, ili barem pojedinačnog hromozoma.
U takvoj šematskoj prezentaciji, strategija totalnog kloniranja izgleda jednostavno. Zapravo, nailazi na ozbiljne poteškoće povezane s potrebom za dobivanjem ogromnog broja klonova (općenito je prihvaćeno da se genom ili njegova regija koja se proučava moraju preklapati klonovima najmanje 10 puta), ogromnom količinom sekvenciranja i izuzetno složenim rad na pristajanju klonova koji zahtijeva učešće stručnjaka za bioinformatiku. Ozbiljna prepreka potpunom kloniranju su različiti repetitivni segmenti DNK, čiji broj, kao što je već spomenuto, naglo raste kako se veličina genoma povećava. Stoga se strategija totalnog sekvenciranja uglavnom koristi u proučavanju genoma virusa i mikroorganizama, iako se uspješno koristi za proučavanje genoma višećelijskog organizma Drosophila.
Rezultati ukupnog sekvenciranja ovog genoma su "superimirani" na ogroman niz informacija o njegovoj hromozomskoj, genskoj i molekularnoj strukturi dobijenih tokom skoro 100-godišnjeg perioda proučavanja Drosophile. Pa ipak, u pogledu stepena sekvenciranja, genom Drosophila (66% ukupne veličine genoma) je značajno inferioran u odnosu na genom Arabidopsis (92%), uprkos njihovim prilično bliskim veličinama - 180 miliona i 125 miliona baznih parova, respektivno. . Stoga je nedavno predloženo da se nazove mješovita tehnologija, koja je korištena za sekvenciranje genoma Drosophila.
Da bi sekvencionirali genom riže, gore spomenute istraživačke grupe su uzele dvije njegove podvrste, koje se najčešće uzgajaju u azijskim zemljama, - Oriza saliva L. ssp indicaj i Oriza saliva L. sspjaponica. Rezultati njihovih studija se u mnogo čemu poklapaju, ali se u mnogo čemu razlikuju. Tako su predstavnici obe grupe izjavili da su dostigli približno 92-93% preklapanja genoma sa kontigom. Pokazalo se da je oko 42% genoma riže predstavljeno kratkim ponavljanjima DNK koji se sastoje od 20 baznih parova, a većina mobilnih DNK elemenata (transposona) nalazi se u intergenskim regijama. Međutim, podaci o veličini genoma riže značajno se razlikuju.
Za japansku podvrstu, veličina genoma je određena na 466 miliona parova baza, a za indijsku 420 miliona. Razlog za ovo odstupanje nije jasan. To može biti posljedica različitih metodoloških pristupa u određivanju veličine nekodirajućeg dijela genoma, odnosno ne odražava pravo stanje stvari. Ali moguće je da postoji razlika od 15% u veličini proučavanih genoma.
Druga velika razlika otkrivena je u broju pronađenih gena: za japansku podvrstu od 46 022 do 55 615 gena po genomu, a za indijsku od 32 000 do 50 000. Razlog ovog odstupanja nije jasan.
Nepotpunost i nedosljednost primljenih informacija konstatuje se u komentarima na objavljene članke. Takođe izražava nadu da će praznine u poznavanju genoma riže biti eliminirane upoređivanjem podataka "grubog sekvenciranja" s rezultatima detaljnog, hijerarhijskog sekvenciranja koje su sproveli učesnici Međunarodnog projekta genoma riže.
KOMPARATIVNA I FUNKCIONALNA BILJNA GENOMIKA
Dobiveni opsežni podaci, od kojih je polovina (rezultati kineske grupe) javno dostupna, nesumnjivo otvaraju široke perspektive kako za proučavanje genoma riže tako i za biljnu genomiku općenito. Poređenje svojstava genoma Arabidopsis i riže pokazalo je da se većina gena (do 80%) identificiranih u genomu Arabidopsis također nalazi u genomu riže; . Istovremeno, 98% gena čija je primarna struktura utvrđena za druge žitarice pronađeno je u genomu pirinča.
Značajna (gotovo dvostruka) razlika između broja gena u riži i Arabidopsisu je zbunjujuća. Istovremeno, podaci nacrta dekodiranja genoma riže dobiveni korištenjem totalnog sekvenciranja praktično se ne upoređuju sa opsežnim rezultatima proučavanja genoma riže metodom hijerarhijskog kloniranja i sekvenciranja, odnosno onoga što je urađeno u na genom Drosophila nije sprovedena. Stoga ostaje nejasno da li razlika u broju gena kod Arabidopsisa i riže odražava pravo stanje stvari ili se objašnjava razlikom u metodološkim pristupima.
Za razliku od genoma Arabidopsis, podaci o genima blizancima u genomu riže nisu dati. Moguće je da njihova relativna količina može biti veća u riži nego u Arabidopsisu. Ovu mogućnost indirektno potkrepljuju podaci o prisutnosti poliploidnih oblika riže. Više jasnoće po ovom pitanju može se očekivati nakon što se završi Međunarodni projekat genoma riže i dobije detaljna slika primarne strukture DNK ovog genoma. Ozbiljne osnove za takvu nadu daje činjenica da se nakon objavljivanja radova o grubom sekvenciranju genoma riže, broj publikacija o strukturi ovog genoma naglo povećao, a posebno su se pojavile informacije o detaljnom sekvenciranju. njegovih 1 i 4 hromozoma.
Poznavanje, barem približno, broja gena u biljkama je od fundamentalnog značaja za uporednu biljnu genomiku. U početku se vjerovalo da, budući da su sve cvjetnice vrlo bliske jedna drugoj po svojim fenotipskim karakteristikama, njihovi genomi bi također trebali biti slični. A ako proučimo genom Arabidopsis, dobićemo informacije o većini genoma drugih biljaka. Indirektna potvrda ove pretpostavke su rezultati sekvenciranja mišjeg genoma, koji je iznenađujuće blizak ljudskom genomu (oko 30 hiljada gena, od kojih se samo 1 tisuću pokazalo drugačijim).
Može se pretpostaviti da razlog razlika između genoma Arabidopsis i riže leži u njihovoj pripadnosti različitim klasama biljaka - dvosupnicama i jednosupnicama. Da bi se ovo pitanje razjasnilo, vrlo je poželjno poznavati barem grubu primarnu strukturu neke druge monokotiledonske biljke. Najrealniji kandidat mogao bi biti kukuruz, čiji je genom približno jednak ljudskom genomu, ali još uvijek mnogo manji od genoma drugih žitarica. Nutritivna vrijednost kukuruza je dobro poznata.
Ogroman materijal dobiven kao rezultat sekvenciranja genoma Arabidopsis i riže postupno postaje osnova za opsežno proučavanje biljnih genoma korištenjem komparativne genomike. Ovakve studije su od opšteg biološkog značaja, jer omogućavaju da se utvrde glavni principi organizacije biljnog genoma u celini i njihovih pojedinačnih hromozoma, da se identifikuju zajedničke karakteristike strukture gena i njihovih regulatornih regiona i da se razmotre omjer funkcionalno aktivnog (genskog) dijela hromozoma i raznih intergenskih DNK regiona koji ne kodiraju proteine. Komparativna genetika također postaje sve važnija za razvoj ljudske funkcionalne genomike. Za komparativne studije provedeno je sekvencioniranje genoma ribe pahuljice i miša.
Jednako važno je proučavanje pojedinačnih gena odgovornih za sintezu pojedinačnih proteina koji određuju specifične tjelesne funkcije. Upravo u otkrivanju, izolaciji, sekvenciranju i određivanju funkcije pojedinačnih gena leži praktičan, prvenstveno medicinski značaj programa Ljudski genom. Ovu okolnost je prije nekoliko godina uočio J. Watson, koji je naglasio da će program Ljudski genom biti završen tek kada se utvrde funkcije svih ljudskih gena.
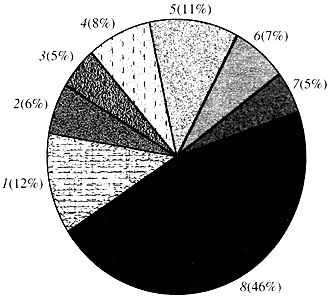
Rice. četiri. Klasifikacija prema funkciji gena Arabidopsis
1 - geni za rast, diobu i sintezu DNK; 2 - geni za sintezu RNK (transkripcija); 3 - geni za sintezu i modifikaciju proteina; 4 - geni za razvoj, starenje i ćelijsku smrt; 5 - geni ćelijskog metabolizma i energetskog metabolizma; 6 - geni međućelijske interakcije i prijenosa signala; 7 - geni za obezbeđivanje drugih ćelijskih procesa; 8 - geni sa nepoznatom funkcijomŠto se tiče funkcije biljnih gena, znamo manje od jedne desetine onoga što znamo o ljudskim genima. Čak i kod Arabidopsis, čiji genom daleko nadmašuje ljudski genom u smislu stepena proučavanja, funkcija skoro polovine njegovih gena ostaje nepoznata (slika 4). U međuvremenu, pored gena uobičajenih za životinje, biljke imaju značajan broj gena koji su specifični samo (ili barem pretežno) za njih. Riječ je o genima uključenim u transport vode i sintezi stanične stijenke, kojih nema kod životinja, genima koji osiguravaju stvaranje i funkcioniranje hloroplasta, fotosintezu, fiksaciju dušika, te sintezu brojnih aromatičnih proizvoda. Ovaj popis se može nastaviti, ali je već jasno s kojim se teškim zadatkom suočava funkcionalna genomika biljaka.
Puno sekvenciranje genoma daje bliske istinitim informacije o ukupnom broju gena u datom organizmu, omogućava da se manje ili više detaljne i pouzdane informacije o njihovoj strukturi smjeste u banke podataka, te olakšava rad na izolaciji i proučavanju pojedinačnih gena. Međutim, sekvenciranje genoma nikako ne znači uspostavljanje funkcije svih gena.
Jedan od pristupa funkcionalne genomike koji najviše obećava zasniva se na identifikaciji radnih gena koji se koriste za transkripciju (čitanje) mRNA. Ovaj pristup, uključujući korištenje moderne tehnologije mikromreža, omogućava simultanu identifikaciju do desetina hiljada funkcionalnih gena. Nedavno je istraživanje biljnih genoma počelo korištenjem ovog pristupa. Za Arabidopsis je bilo moguće dobiti oko 26 hiljada pojedinačnih transkripata, što uvelike olakšava mogućnost određivanja funkcije gotovo svih njegovih gena. U krompiru je bilo moguće identifikovati oko 20.000 radnih gena koji su važni za razumevanje kako procesa rasta i formiranja gomolja, tako i procesa bolesti krompira. Pretpostavlja se da će ovo saznanje povećati otpornost jednog od najvažnijih prehrambenih proizvoda na patogene.
Logičan razvoj funkcionalne genomike bila je proteomika. Ovo novo polje nauke proučava proteom, koji se obično shvata kao kompletan skup proteina u ćeliji u određenom trenutku. Takav skup proteina, koji odražava funkcionalno stanje genoma, stalno se mijenja, dok genom ostaje nepromijenjen.
Proučavanje proteina se dugo koristi za procjenu aktivnosti biljnih genoma. Kao što je poznato, enzimi prisutni u svim biljkama razlikuju se kod pojedinačnih vrsta i varijanti u redoslijedu aminokiselina. Takvi enzimi, sa istom funkcijom, ali različitim slijedom pojedinačnih aminokiselina, nazivaju se izoenzimi. Imaju različita fizičko-hemijska i imunološka svojstva (molekularna težina, naboj), koja se mogu otkriti hromatografijom ili elektroforezom. Dugi niz godina ove metode se uspješno koriste za proučavanje takozvanog genetskog polimorfizma, odnosno razlika između organizama, sorti, populacija, vrsta, posebno pšenice i srodnih oblika žitarica. Nedavno je, međutim, zbog brzog razvoja metoda analize DNK, uključujući sekvenciranje, proučavanje polimorfizma proteina zamijenjeno proučavanjem polimorfizma DNK. Međutim, direktno proučavanje spektra skladišnih proteina (prolamina, glijadina, itd.), koji određuju glavne nutritivne osobine žitarica, ostaje važna i pouzdana metoda za genetičku analizu, selekciju i proizvodnju sjemena poljoprivrednih biljaka.
Poznavanje gena, mehanizama njihove ekspresije i regulacije izuzetno je važno za razvoj biotehnologije i proizvodnju transgenih biljaka. Poznato je da impresivni uspjesi u ovoj oblasti izazivaju dvosmislenu reakciju ekološke i medicinske zajednice. Međutim, postoji područje biljne biotehnologije gdje se ovi strahovi, ako ne i potpuno neutemeljeni, onda, u svakom slučaju, čine malo važnima. Riječ je o stvaranju transgenih industrijskih biljaka koje se ne koriste kao prehrambeni proizvodi. Indija je nedavno pobrala prvi usev transgenog pamuka koji je otporan na brojne bolesti. Postoje informacije o uvođenju posebnih gena koji kodiraju pigmentne proteine u genom pamuka i proizvodnji pamučnih vlakana koja ne zahtijevaju umjetno bojenje. Još jedna industrijska kultura koja može biti predmet efikasnog genetskog inženjeringa je lan. Nedavno se raspravljalo o njegovoj upotrebi kao alternativi pamuku za tekstilne sirovine. Ovaj problem je izuzetno važan za našu zemlju, koja je izgubila sopstvene izvore sirovog pamuka.
PERSPEKTIVE ZA PROUČAVANJE BILJNIH GENOMA
Očigledno je da će se strukturne studije biljnih genoma zasnivati na pristupima i metodama komparativne genomike, koristeći rezultate dešifriranja genoma Arabidopsis i riže kao glavnog materijala. Bitnu ulogu u razvoju komparativne biljne genomike nesumnjivo će imati informacije koje će prije ili kasnije dobiti totalno (grubo) sekvenciranje genoma drugih biljaka. U ovom slučaju, komparativna genomika biljaka će se zasnivati na uspostavljanju genetskih odnosa između pojedinačnih lokusa i hromozoma koji pripadaju različitim genomima. Nećemo se fokusirati toliko na opštu genomiku biljaka koliko na selektivnu genomiku pojedinačnih hromozomskih lokusa. Na primjer, nedavno je pokazano da se gen odgovoran za jarovizaciju nalazi na VRn-AI lokusu heksaploidnog hromozoma 5A pšenice i Hd-6 lokusu rižinog hromozoma 3.
Razvoj ovih studija bit će snažan poticaj za identifikaciju, izolaciju i sekvenciranje mnogih funkcionalno važnih biljnih gena, posebno gena odgovornih za otpornost na bolesti, otpornost na sušu i prilagodljivost različitim uvjetima uzgoja. Sve više će se koristiti funkcionalna genomika, zasnovana na masovnoj detekciji (skriningu) gena koji funkcionišu u biljkama.
Možemo predvidjeti dalje unapređenje hromozomskih tehnologija, prvenstveno metode mikrodisekcije. Njegova upotreba dramatično proširuje mogućnosti genomskog istraživanja bez velikih troškova, kao što je, na primjer, sekvenciranje ukupnog genoma. Metoda lokalizacije na hromozomima biljaka pojedinih gena uz pomoć hibridizacije će se dalje širiti. in situ. Trenutno je njegova primjena ograničena ogromnim brojem ponavljajućih sekvenci u biljnom genomu, a moguće i posebnostima strukturne organizacije biljnih hromozoma.
Kromosomske tehnologije će u doglednoj budućnosti postati od velike važnosti za evolucijsku genomiku biljaka. Ove relativno jeftine tehnologije omogućavaju brzu procjenu intra- i interspecifične varijabilnosti, proučavanje složenih alopoliploidnih genoma tetraploidne i heksaploidne pšenice, tritikala; analizirati evolucijske procese na hromozomskom nivou; istražuju formiranje sintetičkih genoma i unošenje (introgresija) stranog genetskog materijala; identificirati genetske odnose između pojedinačnih hromozoma različitih vrsta.
Za karakterizaciju genoma koristit će se proučavanje kariotipa biljaka klasičnim citogenetskim metodama, obogaćenim molekularno-biološkom analizom i kompjuterskom tehnologijom. Ovo je posebno važno za proučavanje stabilnosti i varijabilnosti kariotipa na nivou ne samo pojedinačnih organizama, već i populacija, varijeteta i vrsta. Konačno, teško je zamisliti kako se broj i spektri kromosomskih preuređivanja (aberacije, mostovi) mogu procijeniti bez upotrebe diferencijalnih metoda bojenja. Ovakve studije su izuzetno obećavajuće za praćenje životne sredine prema stanju biljnog genoma.
U modernoj Rusiji, malo je vjerovatno da će se provesti direktno sekvenciranje biljnih genoma. Takav posao, koji iziskuje velika ulaganja, nepodnošljiv je za našu sadašnju ekonomiju. U međuvremenu, podaci o strukturi genoma Arabidopsis i riže, dobijeni od strane svjetske nauke i dostupni u međunarodnim bankama podataka, dovoljni su za razvoj domaće biljne genomike. Može se predvidjeti proširenje proučavanja biljnih genoma zasnovanih na komparativnim genomskim pristupima za rješavanje specifičnih problema oplemenjivanja i uzgoja biljaka, kao i za proučavanje porijekla različitih biljnih vrsta od velikog ekonomskog značaja.
Može se pretpostaviti da će genomski pristupi kao što je genetska tipizacija (RELF, RAPD, AFLP analize, itd.), koji su prilično pristupačni za naš budžet, imati široku primjenu u domaćoj oplemenjivačkoj praksi i biljnoj proizvodnji. Paralelno sa direktnim metodama za određivanje polimorfizma DNK, pristupi zasnovani na proučavanju polimorfizma proteina, prvenstveno proteina skladištenja žitarica, koristiće se za rješavanje problema genetike i oplemenjivanja biljaka. Kromosomske tehnologije će se široko koristiti. Relativno su jeftini, njihov razvoj zahtijeva prilično umjerena ulaganja. U oblasti proučavanja hromozoma domaća nauka nije inferiorna u odnosu na svet.
Treba naglasiti da je naša nauka dala značajan doprinos nastanku i razvoju biljne genomike [ , ].
Temeljnu ulogu odigrao je N.I. Vavilov (1887-1943).
U molekularnoj biologiji i biljnoj genomici, pionirski doprinos A.N. Belozerski (1905-1972).
U oblasti hromozomskih studija potrebno je istaći rad izuzetnog genetičara S.G. Navašin (1857-1930), koji je prvi otkrio satelitske hromozome u biljkama i dokazao da je moguće razlikovati pojedinačne hromozome prema karakteristikama njihove morfologije.
Još jedan klasik ruske nauke G.A. Levitsky (1878-1942) je detaljno opisao hromozome raži, pšenice, ječma, graška i šećerne repe, uveo pojam "kariotip" u nauku i razvio doktrinu o tome.
Savremeni stručnjaci, oslanjajući se na dostignuća svjetske nauke, mogu dati značajan doprinos daljem razvoju biljne genetike i genomike.
Autor se od srca zahvaljuje akademiku Yu.P. Altukhovu za kritičku raspravu o članku i vrijedne savjete.Rad tima na čelu sa autorom članka podržala je Ruska fondacija za osnovna istraživanja (grantovi br. 99-04-48832; 00-04-49036; 00-04-81086), Program predsjednika Ruske Federacije za podršku naučnim školama (grantovi br. 00-115 -97833 i NSh-1794.2003.4) i Program Ruske akademije nauka "Molekularno genetski i hromozomski markeri u razvoju savremenih metoda oplemenjivanja i proizvodnje sjemena" .
LITERATURA
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Uvod u biljnu genomiku // Molekularna biologija. 2001. V. 35. S. 339-348. 2. Pen E. Bonanza za biljnu genomiku // Science. 1998. V. 282. P. 652-654. 3. Biljna genomika, Proc. Natl. Akad. sci. SAD. 1998. V. 95. P. 1962-2032. 4. Cartel N.A. i sl. Genetika. Encyclopedic Dictionary. Minsk: Tehnologija, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferencijacija genoma u Aegilopsu. 1. Distribucija visoko repetitivnih sekvenci DNK na hromozomima diploidnih vrsta, Genom. 1996. V. 39. P. 293-306. Povijest analize hromozoma // Biol. membrane. 2001. T. 18. S. 164-172.
Geni za skakanje
Sredinom prošlog stoljeća američka istraživačica Barbara McClintock otkrila je nevjerovatne gene u kukuruzu koji mogu samostalno mijenjati svoj položaj na hromozomima. Sada se nazivaju "geni za skakanje" ili prenosivi (pokretni) elementi. Ovo otkriće dugo nije bilo prepoznato, smatrajući da su pokretni elementi jedinstvena pojava, karakteristična samo za kukuruz. Međutim, za ovo otkriće je 1983. godine nagrađen McClintock nobelova nagrada Danas su geni za skakanje pronađeni u gotovo svim proučavanim životinjskim i biljnim vrstama.
Odakle potiču geni za skakanje, šta rade u ćeliji, ima li koristi od njih? Zašto, uz genetski zdrave roditelje, porodica voćne mušice Drosophila, zbog gena za skakanje, može sa velikom frekvencijom proizvesti mutantno potomstvo ili čak biti potpuno bez djece? Koja je uloga gena za skok u evoluciji?
Mora se reći da su geni koji osiguravaju funkcioniranje stanica smješteni na hromozomima određenim redoslijedom. Zahvaljujući tome, za mnoge vrste jednoćelijskih i višećelijskih organizama bilo je moguće izgraditi tzv. genetske karte. Međutim, između gena postoji red veličine više genetskog materijala nego u njima samima! Kakvu ulogu igra ovaj "balastni" dio DNK nije do kraja utvrđeno, ali se tu najčešće nalaze mobilni elementi, koji ne samo da se sami pokreću, već mogu sa sobom ponijeti i susjedne fragmente DNK.

Odakle potiču geni skakača? Vjeruje se da barem neki od njih potječu od virusa, budući da neki mobilni elementi mogu formirati virusne čestice (na primjer, ciganski mobilni element u voćnoj mušici Drosophila melanogaster). Neki prenosivi elementi pojavljuju se u genomu tzv horizontalni transfer od drugih vrsta. Na primjer, utvrđeno je da mobilni skitnica-element (u prevodu na ruski zove se skitnica) Drosophila melanogaster više puta uveden u genom ove vrste. Postoji verzija da neki regulatorni regioni DNK takođe mogu imati autonomiju i sklonost "skitnji".
korisni balast
S druge strane, većina gena za skakanje, uprkos imenu, ponaša se tiho, iako čine petinu ukupnog genetskog materijala. Drosophila melanogaster ili skoro polovina ljudskog genoma.
Redundantnost DNK, koja je gore spomenuta, ima svoj plus: balastna DNK (uključujući pasivne mobilne elemente) preuzima udarac ako se strana DNK unese u genom. Vjerovatnoća da će se novi element umetnuti u koristan gen i time poremetiti njegov rad je smanjena ako je DNK mnogo više nego značajan.
Neka redundancija DNK je korisna na isti način kao i "suvišak" slova u riječima: pišemo "Marija Ivanovna" i kažemo "Marivana". Neka slova se neminovno gube, ali značenje ostaje. Isti princip funkcioniše i na nivou značaja pojedinačnih aminokiselina u molekulu protein-enzim: samo je redosled aminokiselina koji formira aktivni centar strogo konzervativan. Dakle, na različitim nivoima, redundantnost se ispostavlja kao neka vrsta bafera koji obezbjeđuje sigurnosnu marginu za sistem. Tako mobilni elementi koji su izgubili mobilnost nisu beskorisni za genom. Kako kažu, "čak i čuperak vune od tanke ovce", iako bi, možda, ovdje bolje pristajala druga poslovica - "svako u nizu".

Mobilni elementi koji su zadržali sposobnost skakanja kreću se duž hromozoma Drosophila na frekvenciji od 10–2–10–5 po genu po generaciji, ovisno o vrsti elementa, genetskoj pozadini i spoljni uslovi. To znači da jedan od stotinu gena za skok u ćeliji može promijeniti svoju poziciju nakon sljedeće diobe ćelije. Kao rezultat, nakon nekoliko generacija, distribucija prijenosnih elemenata duž hromozoma može se vrlo značajno promijeniti.
Pogodno je proučavati takvu raspodjelu na politenskim (multifilamentnim) hromozomima iz pljuvačnih žlijezda larvi Drosophila. Ovi hromozomi su mnogo puta deblji od normalnih, što ih čini mnogo lakšim za ispitivanje pod mikroskopom. Kako nastaju ovi hromozomi? U ćelijama pljuvačnih žlijezda, DNK svakog od hromozoma se umnožava, kao u normalnoj diobi ćelije, ali se sama stanica ne dijeli. Kao rezultat toga, broj ćelija u žlijezdi se ne mijenja, ali se u 10-11 ciklusa nakuplja nekoliko hiljada identičnih DNK lanaca u svakom hromozomu.

Djelomično zbog politenskih hromozoma, geni za skakanje kod Drosophile su bolje proučeni nego kod drugih metazoa. Kao rezultat ovih istraživanja, pokazalo se da je čak i unutar iste populacije Drosophila teško pronaći dvije individue koje imaju hromozome sa istom raspodjelom mobilnih elemenata. Nije slučajno što se vjeruje da je većina spontanih mutacija kod Drosophile uzrokovana kretanjem ovih "skokova".
Posljedice mogu biti različite...
Na osnovu njihovog djelovanja na genom, aktivni prenosivi elementi mogu se podijeliti u nekoliko grupa. Neki od njih obavljaju funkcije koje su izuzetno važne i korisne za genom. Na primjer, telomerni DNK, koja se nalazi na krajevima hromozoma, kod Drosophile se sastoji samo od posebnih mobilnih elemenata. Ova DNK je izuzetno važna - njen gubitak povlači gubitak cijelog hromozoma u procesu diobe ćelije, što dovodi do smrti ćelije.
Ostali mobilni elementi su direktne "štetočine". Barem se smatraju takvima ovog trenutka. Na primjer, prenosivi elementi klase R2 mogu se posebno uvesti u gene člankonožaca koji kodiraju jedan od proteina ribozoma - stanične "tvornice" za sintezu proteina. Pojedinci s takvim poremećajima preživljavaju samo zato što je samo dio mnogih gena koji kodiraju ove proteine oštećen u genomu.

Postoje i takvi mobilni elementi koji se kreću samo u reproduktivnim tkivima koji proizvode zametne stanice. To se objašnjava činjenicom da u različitim tkivima isti mobilni element može proizvesti različite dužine i funkcije molekula protein-enzim neophodnih za kretanje.
Primjer potonjeg je P-element Drosophila melanogaster, koja je u svoje prirodne populacije dospjela horizontalnim prijenosom s druge vrste Drosophila prije ne više od stotinu godina. Međutim, trenutno na Zemlji jedva da postoji stanovništvo. Drosophila melanogaster, u kojem ne bi bilo P-elementa. Istovremeno, treba napomenuti da je većina njegovih kopija neispravna, štoviše, ista verzija kvara pronađena je gotovo posvuda. Uloga potonjeg u genomu je osebujna: "netolerantna" je prema svojim bližnjima i igra ulogu represora, blokirajući njihovo kretanje. Dakle, zaštita genoma Drosophila od skokova "vanzemaljaca" može se djelomično provesti vlastitim derivatima.
Glavna stvar je odabrati prave roditelje!
Većina skokova mobilnih elemenata ne utiče izgled Drosophila, jer pada na balastni DNK, ali postoje i druge situacije kada se njihova aktivnost dramatično povećava.

Čudno je da je najmoćniji faktor koji indukuje kretanje gena za skakanje loše roditeljstvo. Na primjer, šta se dešava ako ukrstite ženke iz laboratorijske populacije Drosophila melanogaster, koji nemaju P-element (jer su njihovi preci uhvaćeni iz prirode prije stotinjak godina), a mužjaci nose P-element? Kod hibrida se zbog brzog kretanja mobilnog elementa može pojaviti veliki broj različitih genetskih poremećaja. Ovaj fenomen, nazvan hibridna disgeneza, uzrokovan je odsustvom represora u citoplazmi majke koji sprječava kretanje mobilnog elementa.
Dakle, ako mladoženja iz populacije A i nevjeste iz populacije B mogu stvoriti velike porodice, onda suprotno nije uvijek tačno. Porodica genetski zdravih roditelja može proizvesti veliki broj mutantnih ili sterilnih potomaka, ili čak ostati bez djece ako otac i majka imaju različit skup mobilnih elemenata u genomu. Posebno mnogo kršenja se pojavljuju ako se eksperiment provodi na temperaturi od 29 ° C. Utjecaj vanjskih faktora, koji se nadovezuju na genetsku pozadinu, pojačava učinak neusklađenosti genoma, iako sami ovi faktori (čak i jonizujuće zračenje) sami po sebi nisu sposobni izazivanja tako masovnog kretanja mobilnih elemenata.
Slični događaji u Drosophila melanogaster može se desiti uz učešće drugih porodica mobilnih elemenata.
"Mobilna" evolucija
Ćelijski genom se može posmatrati kao svojevrsni ekosistem stalnih i privremenih članova, gde susedi ne samo da koegzistiraju, već i međusobno komuniciraju. Interakcija gena domaćina sa prenosivim elementima je još uvijek slabo shvaćena, ali se mogu navesti mnogi rezultati - od smrti organizma u slučaju oštećenja važnog gena do obnove prethodno oštećenih funkcija.
Dešava se da sami geni za skakanje međusobno djeluju. Dakle, poznat je fenomen koji liči na imunitet, kada se mobilni element ne može uvesti u neposrednu blizinu postojećeg. Međutim, nisu svi mobilni elementi tako delikatni: na primjer, R-elementi se lako mogu ugraditi jedan u drugi i izbaciti svoju braću iz igre.

Osim toga, postoji svojevrsna samoregulacija broja transpobilnih elemenata u genomu. Činjenica je da mobilni elementi mogu međusobno razmjenjivati homologne regije - ovaj proces se zove rekombinacija. Kao rezultat takve interakcije, mobilni elementi mogu, ovisno o svojoj orijentaciji, izgubiti ( brisanje) ili proširiti ( inverzija) fragmenti DNK domaćina koji se nalaze između njih. Ako se izgubi značajan dio hromozoma, genom će umrijeti. U slučaju inverzije ili male delecije stvara se hromozomska raznolikost, što se smatra neophodnim uslovom za evoluciju.
Ako dođe do rekombinacija između mobilnih elemenata koji se nalaze na različitim kromosomima, tada nastaju kromosomski preustroji, koji tijekom narednih dioba stanica mogu dovesti do neravnoteže u genomu. A neuravnotežen genom, kao i neuravnotežen budžet, vrlo je loše podijeljen. Dakle, smrt neuspješnih genoma je jedan od razloga zašto aktivni prenosivi elementi ne preplavljuju hromozome bez ograničenja.
Postavlja se prirodno pitanje: koliko je značajan doprinos mobilnih elemenata evoluciji? Prvo, većina prenosivih elemenata se uvodi, grubo rečeno, tamo gdje mora, uslijed čega mogu oštetiti ili promijeniti strukturu ili regulaciju gena u koji su uneseni. Tada prirodna selekcija odbacuje neuspješne opcije, a uspješne opcije s prilagodljivim svojstvima se fiksiraju.

Ako se posljedice uvođenja prijenosnog elementa pokažu neutralnim, tada se ova varijanta može sačuvati u populaciji, pružajući određenu raznolikost u strukturi gena. Ovo može biti korisno kada nepovoljni uslovi. Teoretski, tokom masovnog kretanja mobilnih elemenata, mutacije se mogu pojaviti u više gena istovremeno, što može biti vrlo korisno u slučaju nagle promjene uslova postojanja.
Dakle, da rezimiramo: postoji mnogo mobilnih elemenata u genomu i oni su različiti; mogu komunicirati i jedni s drugima i sa genima domaćina; može biti štetna i nezamjenjiva. Nestabilnost genoma uzrokovana kretanjem mobilnih elemenata može završiti tragedijom za pojedinca, ali sposobnost brze promjene neophodan je uvjet za opstanak populacije ili vrste. To stvara raznolikost, koja je osnova za prirodnu selekciju i kasniju evolucijsku transformaciju.
Možete povući neku analogiju između skačućih gena i imigranata: neki imigranti ili njihovi potomci postaju ravnopravni građani, drugima se daje boravišna dozvola, a treći - oni koji se ne pridržavaju zakona - bivaju deportovani ili zatvoreni. A masovne migracije naroda mogu brzo promijeniti i samu državu.
Književnost
Ratner V. A., Vasilyeva L. A. Indukcija transpozicije mobilnih genetskih elemenata stresnim utjecajima. Ruski povez. 2000.
Gvozdev V. A. Pokretna eukariotska DNK // Soros Educational Journal. 1998. br. 8.





